

Tight junction pore and leak pathways: a dynamic duo
- PMID: 20936941
- PMCID: PMC4655434
- DOI: 10.1146/annurev-physiol-012110-142150
Tight junction pore and leak pathways: a dynamic duo
Abstract
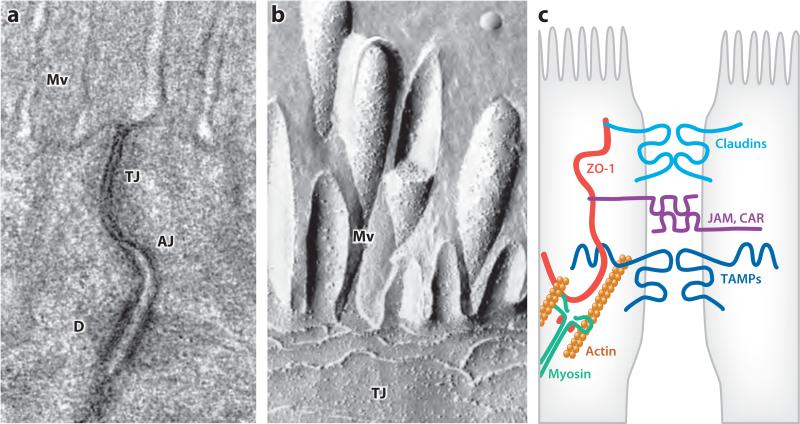
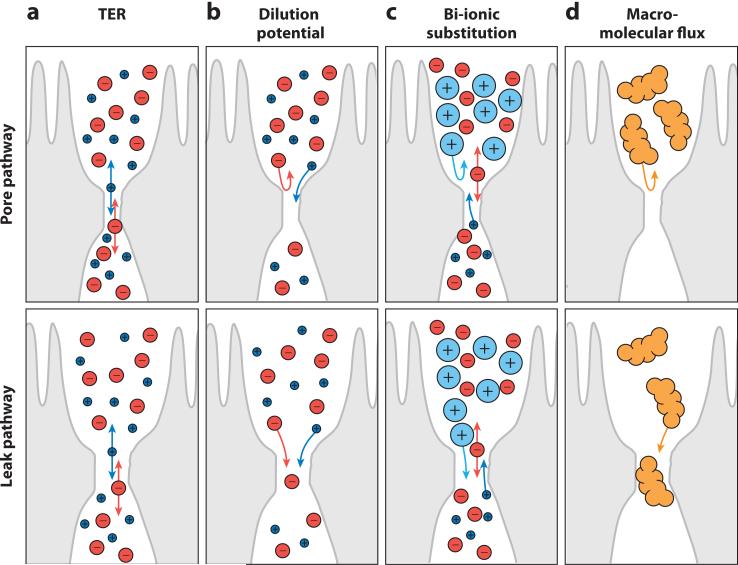
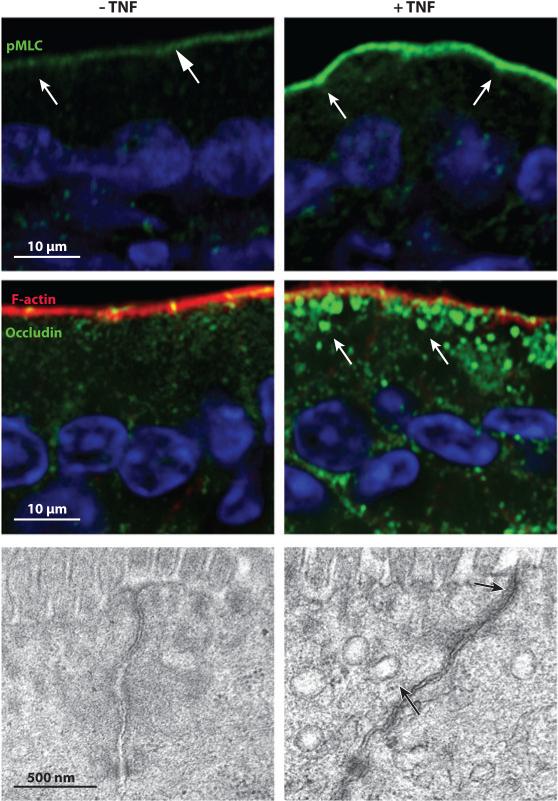
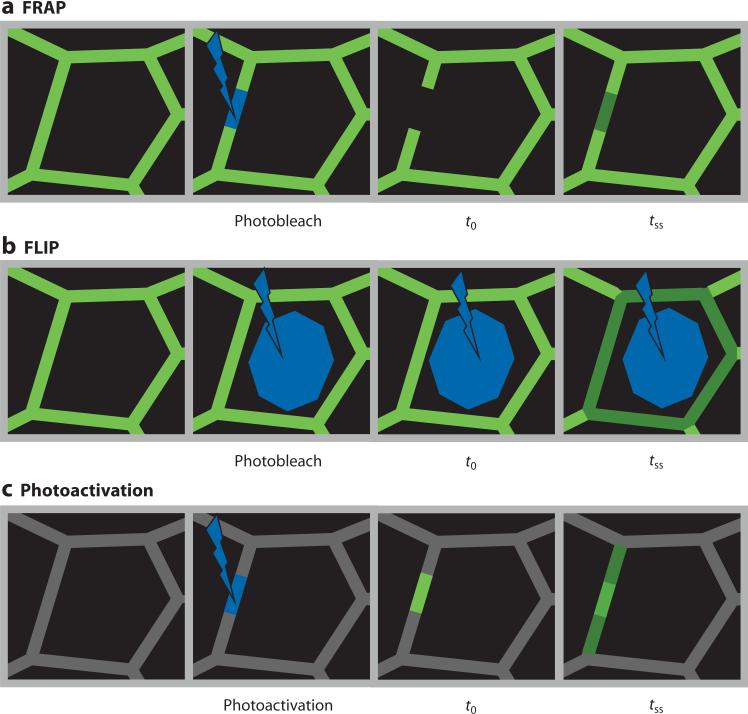
Tissue barriers that restrict passage of liquids, ions, and larger solutes are essential for the development of multicellular organisms. In simple organisms this allows distinct cell types to interface with the external environment. In more complex species, the diversity of cell types capable of forming barriers increases dramatically. Although the plasma membranes of these barrier-forming cells prevent flux of most hydrophilic solutes, the paracellular, or shunt, pathway between cells must also be sealed. This function is accomplished in vertebrates by the zonula occludens, or tight junction. The tight junction barrier is not absolute but is selectively permeable and is able to discriminate between solutes on the basis of size and charge. Many tight junction components have been identified over the past 20 years, and recent progress has provided new insights into the proteins and interactions that regulate structure and function. This review presents these data in a historical context and proposes an integrated model in which dynamic regulation of tight junction protein interactions determines barrier function.
Figures
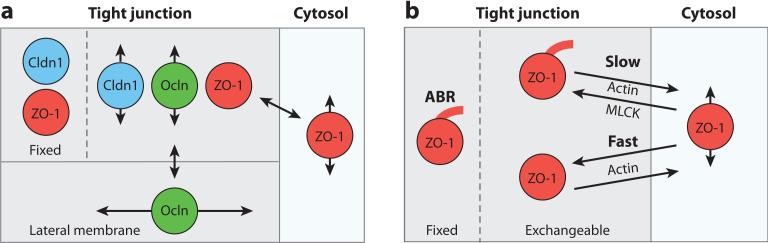
References
-
- Foskett JK, Machen TE. Vibrating probe analysis of teleost opercular epithelium: correlation between active transport and leak pathways of individual chloride cells. J. Membr. Biol. 1985;85:25–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01DK067887/DK/NIDDK NIH HHS/United States
- P30CA14599/CA/NCI NIH HHS/United States
- UL1 RR024999/RR/NCRR NIH HHS/United States
- P30 CA014599/CA/NCI NIH HHS/United States
- R01DK68271/DK/NIDDK NIH HHS/United States
- T32 GM007281/GM/NIGMS NIH HHS/United States
- R01 DK061931/DK/NIDDK NIH HHS/United States
- F32 DK082134/DK/NIDDK NIH HHS/United States
- 5T32GM007281/GM/NIGMS NIH HHS/United States
- P30DK042086/DK/NIDDK NIH HHS/United States
- UL1RR024999/RR/NCRR NIH HHS/United States
- F32DK082134/DK/NIDDK NIH HHS/United States
- P01 DK067887/DK/NIDDK NIH HHS/United States
- P30 DK042086/DK/NIDDK NIH HHS/United States
- R01 DK068271/DK/NIDDK NIH HHS/United States
- K08 DK088953/DK/NIDDK NIH HHS/United States
- R01DK61931/DK/NIDDK NIH HHS/United States
- K08DK088953/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
