

Molecular mechanism of allosteric modification of voltage-dependent sodium channels by local anesthetics
- PMID: 20937693
- PMCID: PMC2964522
- DOI: 10.1085/jgp.201010438
Molecular mechanism of allosteric modification of voltage-dependent sodium channels by local anesthetics
Abstract
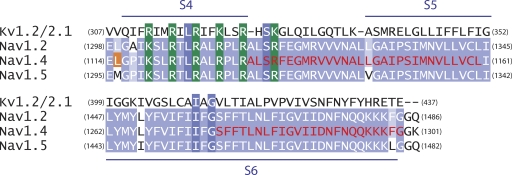
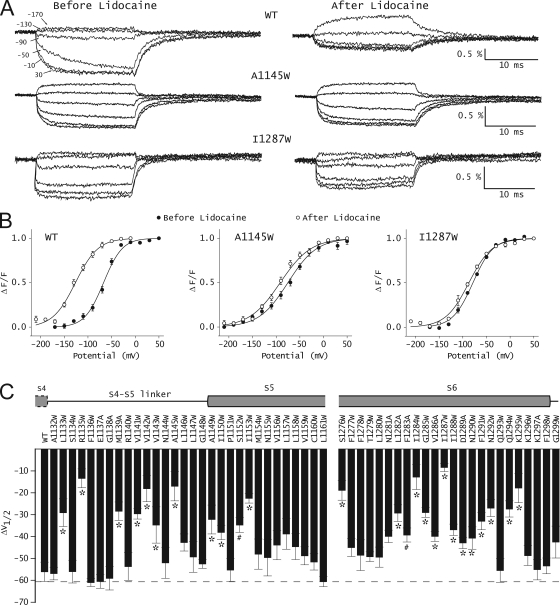
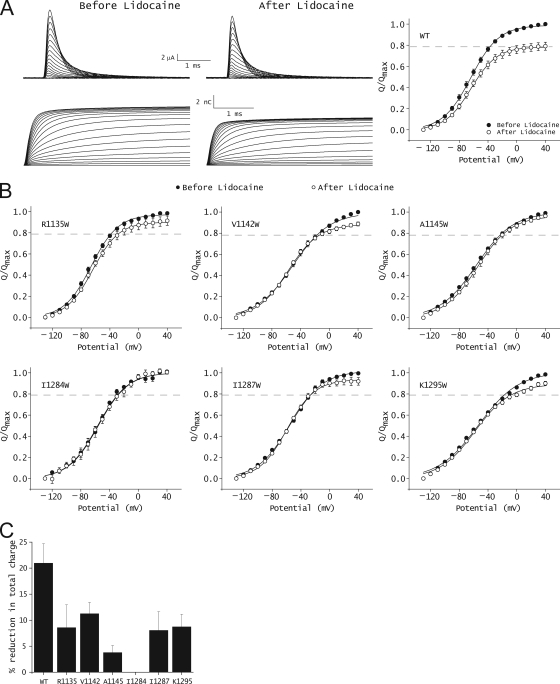
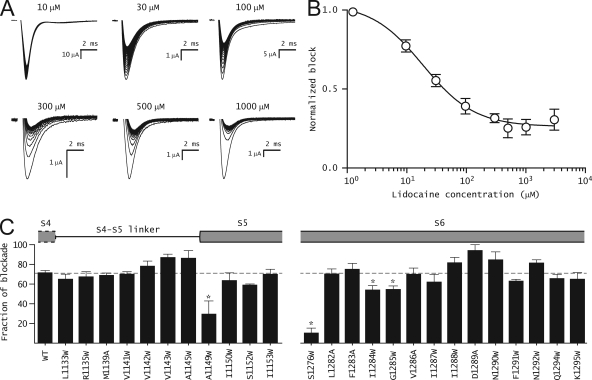
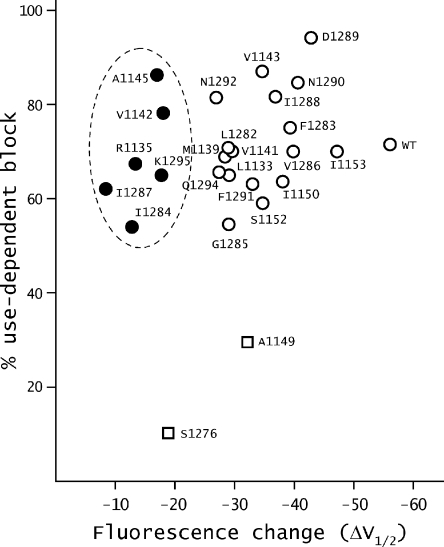
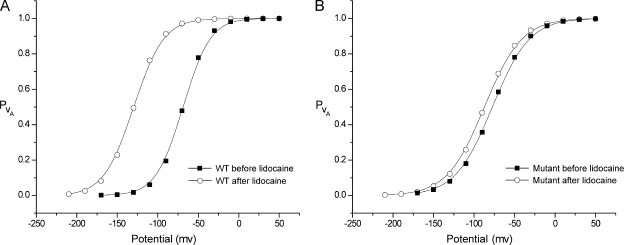
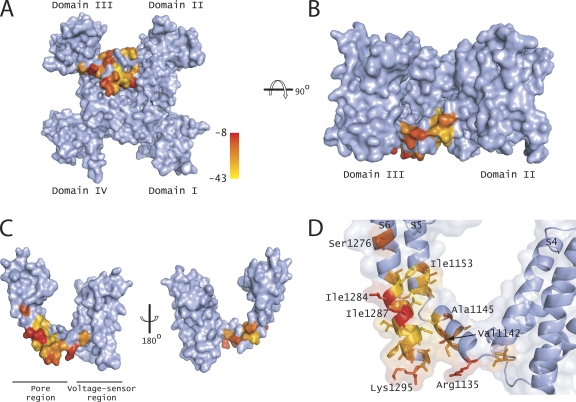
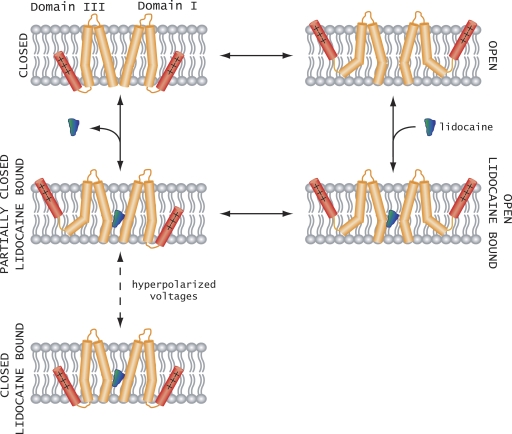
The hallmark of many intracellular pore blockers such as tetra-alkylammonium compounds and local anesthetics is their ability to allosterically modify the movement of the voltage sensors in voltage-dependent ion channels. For instance, the voltage sensor of domain III is specifically stabilized in the activated state when sodium currents are blocked by local anesthetics. The molecular mechanism underlying this long-range interaction between the blocker-binding site in the pore and voltage sensors remains poorly understood. Here, using scanning mutagenesis in combination with voltage clamp fluorimetry, we systematically evaluate the role of the internal gating interface of domain III of the sodium channel. We find that several mutations in the S4-S5 linker and S5 and S6 helices dramatically reduce the stabilizing effect of lidocaine on the activation of domain III voltage sensor without significantly altering use-dependent block at saturating drug concentrations. In the wild-type skeletal muscle sodium channel, local anesthetic block is accompanied by a 21% reduction in the total gating charge. In contrast, point mutations in this critical intracellular region reduce this charge modification by local anesthetics. Our analysis of a simple model suggests that these mutations in the gating interface are likely to disrupt the various coupling interactions between the voltage sensor and the pore of the sodium channel. These findings provide a molecular framework for understanding the mechanisms underlying allosteric interactions between a drug-binding site and voltage sensors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
