

Functional central spindle assembly requires de novo microtubule generation in the interchromosomal region during anaphase
- PMID: 20937700
- PMCID: PMC2958471
- DOI: 10.1083/jcb.201004150
Functional central spindle assembly requires de novo microtubule generation in the interchromosomal region during anaphase
Abstract
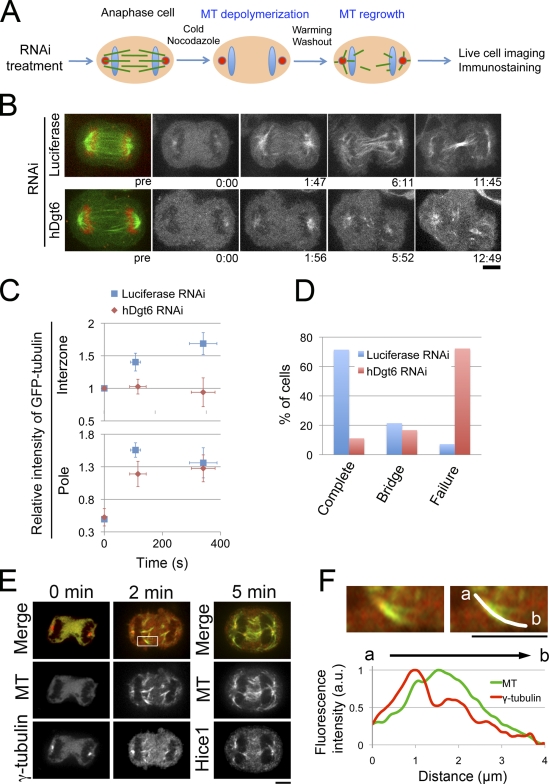
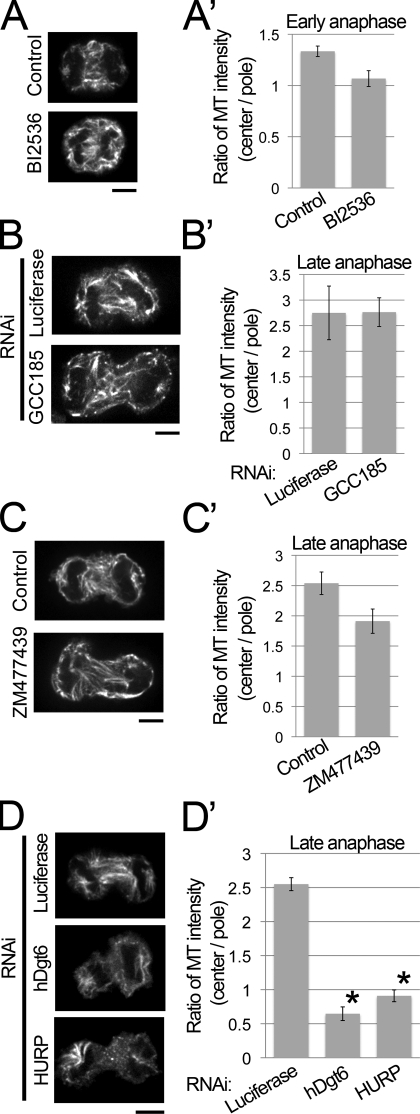
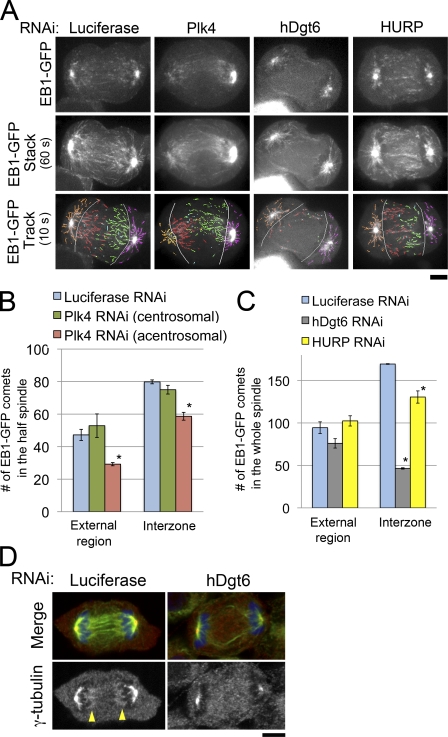
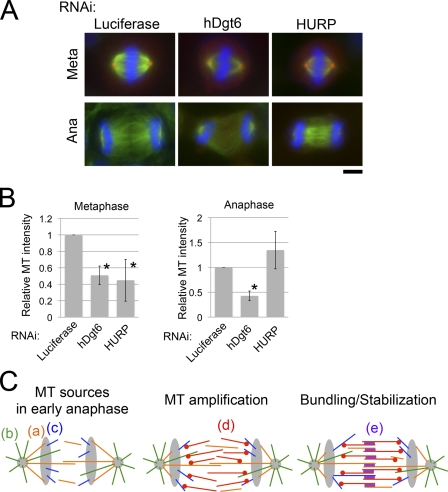
The central spindle forms between segregating chromosomes during anaphase and is required for cytokinesis. Although anaphase-specific bundling and stabilization of interpolar microtubules (MTs) contribute to formation of the central spindle, it remains largely unknown how these MTs are prepared. Using live imaging of MT plus ends and an MT depolymerization and regrowth assay, we show that de novo MT generation in the interchromosomal region during anaphase is important for central spindle formation in human cells. Generation of interchromosomal MTs and subsequent formation of the central spindle occur independently of preanaphase MTs or centrosomal MT nucleation but require augmin, a protein complex implicated in nucleation of noncentrosomal MTs during preanaphase. MTs generated in a hepatoma up-regulated protein (HURP)-dependent manner during anaphase also contribute to central spindle formation redundantly with preanaphase MTs. Based on these results, a new model for central spindle assembly is proposed.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
