

The early secretory pathway contributes to the growth of the Coxiella-replicative niche
- PMID: 20937765
- PMCID: PMC3019900
- DOI: 10.1128/IAI.00688-10
The early secretory pathway contributes to the growth of the Coxiella-replicative niche
Abstract
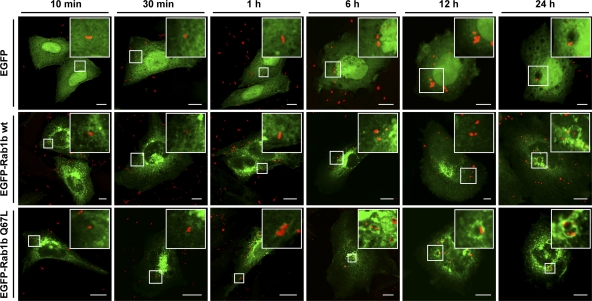
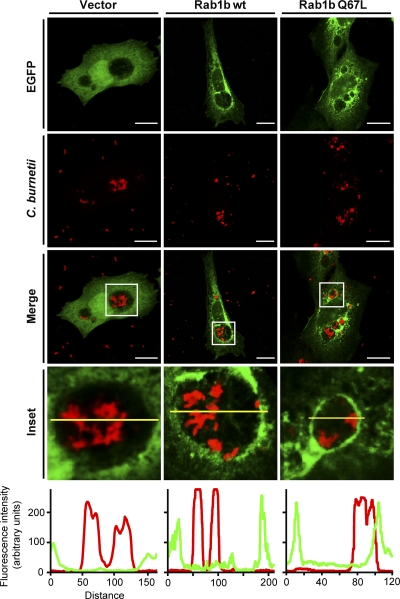
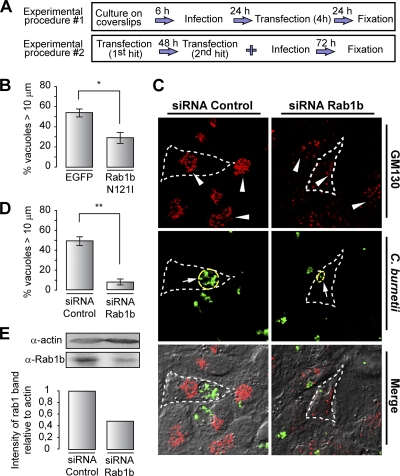
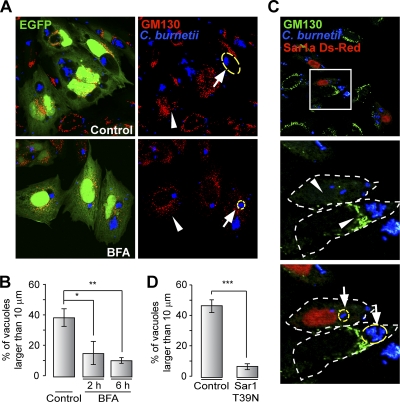
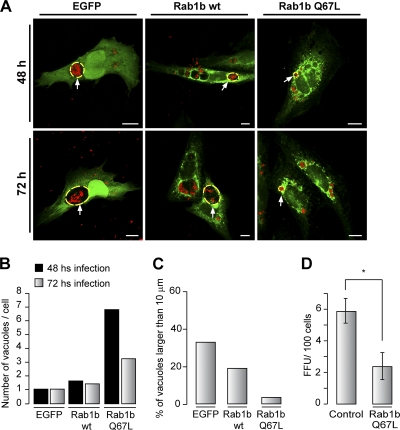
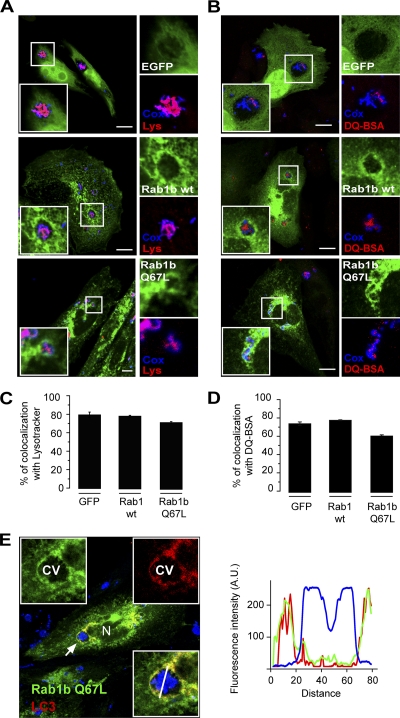
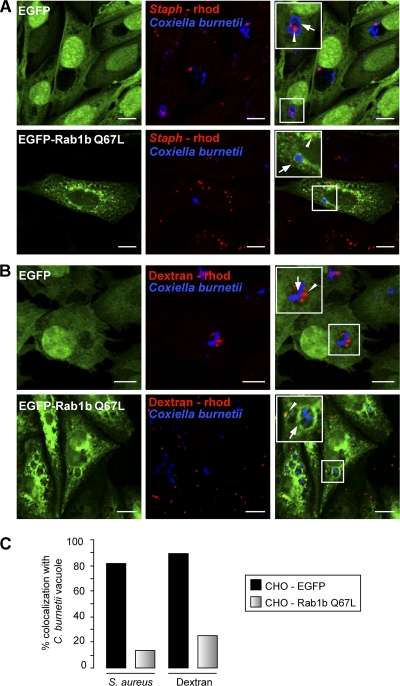
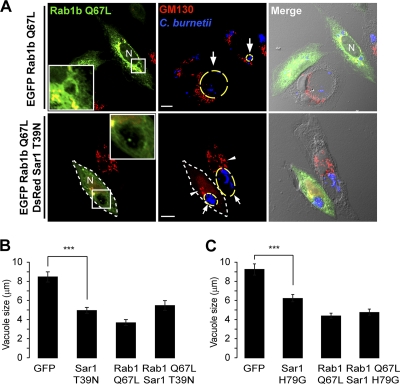
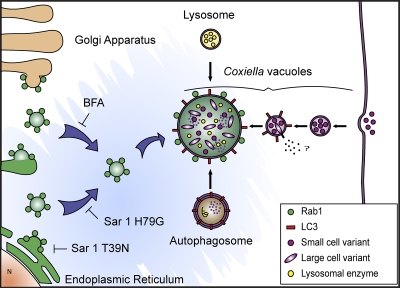
Coxiella burnetii is a Gram-negative obligate intracellular bacterium. After internalization, this bacterium replicates in a large parasitophorous vacuole that has features of both phagolysosomes and autophagosomal compartments. We have previously demonstrated that early after internalization Coxiella phagosomes interact with both the endocytic and the autophagic pathways. In this report, we present evidence that the Coxiella-replicative vacuoles (CRVs) also interact with the secretory pathway. Rab1b is a small GTPase responsible for the anterograde transport between the endoplasmic reticulum and the Golgi apparatus. We present evidence that Rab1b is recruited to the CRV at later infection times (i.e., after 6 h of infection). Interestingly, knockdown of Rab1b altered vacuole growth, indicating that this protein was required for the proper biogenesis of the CRV. In addition, overexpression of the active GTPase-defective mutant (GFP-Rab1b Q67L) affected the development of the Coxiella-replicative compartment inhibiting bacterial growth. On the other hand, disruption of the secretory pathway by brefeldin A treatment or by overexpression of Sar1 T39N, a defective dominant-negative mutant of Sar1, affected the typical spaciousness of the CRVs. Taken together, our results show for the first time that the Coxiella-replicative niche also intercepts the early secretory pathway.
Figures
References
-
- Allan, B. B., B. D. Moyer, and W. E. Balch. 2000. Rab1 recruitment of p115 into a cis-SNARE complex: programming budding COPII vesicles for fusion. Science 289:444-448. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
