

Temperature and Mg2+ sensing by a novel PhoP-PhoQ two-component system for regulation of virulence in Edwardsiella tarda
- PMID: 20937832
- PMCID: PMC2998078
- DOI: 10.1074/jbc.M110.179150
Temperature and Mg2+ sensing by a novel PhoP-PhoQ two-component system for regulation of virulence in Edwardsiella tarda
Abstract
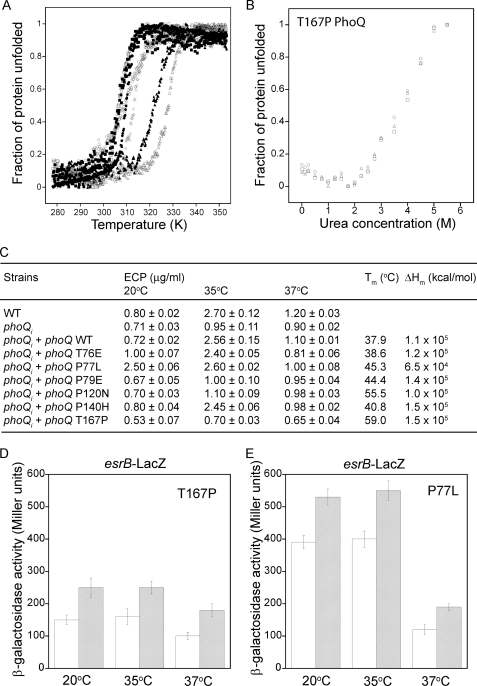
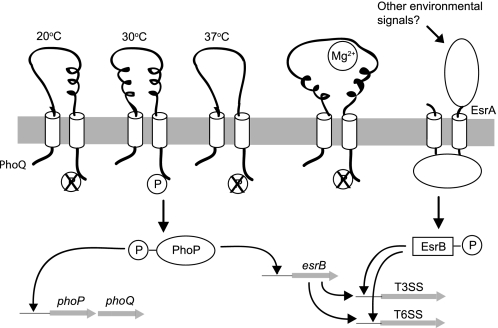
The PhoP-PhoQ two-component system is commonly used by bacteria to sense environmental factors. Here we show that the PhoP-PhoQ system of Edwardsiella tarda detects changes in environmental temperature and Mg(2+) concentration as well as regulates the type III and VI secretion systems through direct activation of esrB. Protein secretion is activated from 23 to 35 °C or at low Mg(2+) concentrations, but it is suppressed at or below 20 °C, at or above 37 °C, or at high Mg(2+) concentrations. The effects of temperature and Mg(2+) concentration are additive. The PhoQ sensor domain has a low T(m) of 37.9 °C, and it detects temperatures through a conformational change of its secondary structure. Mutation of specific Pro or Thr residues increased the stability of the PhoQ sensor drastically, altering its temperature-sensing ability. The PhoQ sensor detects Mg(2+) concentration through the direct binding of Mg(2+) to a cluster of acidic residues (DDDSAD) and through changes that likely affect its tertiary structure. Here, we describe for the first time the use of PhoP-PhoQ as a temperature sensor for bacterial virulence control.
Figures





References
-
- Janda J. M., Abbott S. L. (1993) Clin. Infect. Dis. 17, 742–748 - PubMed
-
- Thune R. L., Stanley L. A., Cooper R. K. (1993) Annu. Rev. Fish Dis. 3, 37–68
-
- Srinivasa Rao P. S., Yamada Y., Tan Y. P., Leung K. Y. (2004) Mol. Microbiol. 53, 573–586 - PubMed
-
- Tan Y. P., Zheng J., Tung S. L., Rosenshine I., Leung K. Y. (2005) Microbiology 151, 2301–2313 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
