

The evolution of vertebrate tetraspanins: gene loss, retention, and massive positive selection after whole genome duplications
- PMID: 20939927
- PMCID: PMC2965184
- DOI: 10.1186/1471-2148-10-306
The evolution of vertebrate tetraspanins: gene loss, retention, and massive positive selection after whole genome duplications
Abstract
Background: The vertebrate tetraspanin family has many features which make it suitable for preserving the imprint of ancient sequence evolution and amenable for phylogenomic analysis. So we believe that an in-depth analysis of the tetraspanin evolution not only provides more complete understanding of tetraspanin biology, but offers new insights into the influence of the two rounds of whole genome duplication (2R-WGD) at the origin of vertebrates.
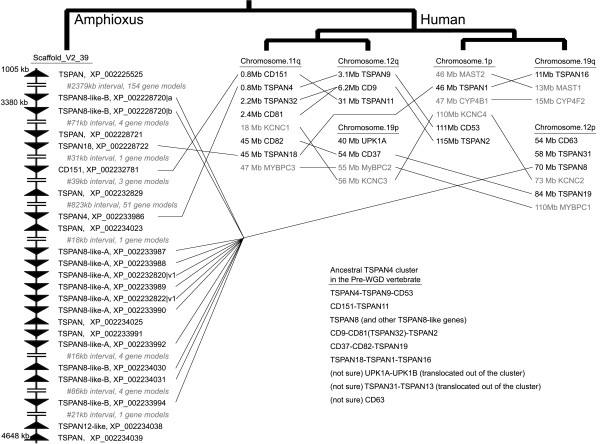
Results: A detailed phylogeny of vertebrate tetraspanins was constructed by using multiple lines of information, including sequence-based phylogenetics, key structural features, intron configuration and genomic synteny. In particular, a total of 38 modern tetraspanin ortholog lineages in bony vertebrates have been identified and subsequently classified into 17 ancestral lineages existing before 2R-WGD. Based on this phylogeny, we found that the ohnolog retention rate of tetraspanins after 2R-WGD was three times as the average (a rate similar to those of transcription factors and protein kinases). This high rate didn't increase the tetrapanin family size, but changed the family composition, possibly by displacing vertebrate-specific gene lineages with the lineages conserved across deuterostomes. We also found that the period from 2R-WGD to recent time is controlled by gene losses. Meanwhile, positive selection has been detected on 80% of the branches right after 2R-WGDs, which declines significantly on both magnitude and extensity on the following speciation branches. Notably, the loss of mammalian RDS2 is accompanied by strong positive selection on mammalian ROM1, possibly due to gene loss-induced compensatory evolution.
Conclusions: First, different from transcription factors and kinases, high duplicate retention rate after 2R-WGD didn't increase the tetraspanin family size but just reshaped the family composition. Second, the evolution of tetraspanins right after 2R-WGD had been impacted by a massive wave of gene loss and positive selection on coding sequences. Third, the lingering effect of 2R-WGD on tetraspanin gene loss and positive selection might last for 300-400 million years.
Figures




Similar articles
-
OHNOLOGS v2: a comprehensive resource for the genes retained from whole genome duplication in vertebrates.Nucleic Acids Res. 2020 Jan 8;48(D1):D724-D730. doi: 10.1093/nar/gkz909. Nucleic Acids Res. 2020. PMID: 31612943 Free PMC article.
-
2R and remodeling of vertebrate signal transduction engine.BMC Biol. 2010 Dec 13;8:146. doi: 10.1186/1741-7007-8-146. BMC Biol. 2010. PMID: 21144020 Free PMC article.
-
Whole Genome Duplications Shaped the Receptor Tyrosine Kinase Repertoire of Jawed Vertebrates.Genome Biol Evol. 2016 Jun 3;8(5):1600-13. doi: 10.1093/gbe/evw103. Genome Biol Evol. 2016. PMID: 27260203 Free PMC article.
-
Impact of gene gains, losses and duplication modes on the origin and diversification of vertebrates.Semin Cell Dev Biol. 2013 Feb;24(2):83-94. doi: 10.1016/j.semcdb.2012.12.008. Epub 2013 Jan 3. Semin Cell Dev Biol. 2013. PMID: 23291262 Review.
-
Hox gene clusters of early vertebrates: do they serve as reliable markers for genome evolution?Genomics Proteomics Bioinformatics. 2011 Jun;9(3):97-103. doi: 10.1016/S1672-0229(11)60012-0. Genomics Proteomics Bioinformatics. 2011. PMID: 21802046 Free PMC article. Review.
Cited by
-
Fish lateral line innovation: insights into the evolutionary genomic dynamics of a unique mechanosensory organ.Mol Biol Evol. 2012 Dec;29(12):3887-98. doi: 10.1093/molbev/mss194. Epub 2012 Jul 27. Mol Biol Evol. 2012. PMID: 22844072 Free PMC article.
-
Tetraspanin Cd9b and Cxcl12a/Cxcr4b have a synergistic effect on the control of collective cell migration.PLoS One. 2021 Nov 30;16(11):e0260372. doi: 10.1371/journal.pone.0260372. eCollection 2021. PLoS One. 2021. PMID: 34847198 Free PMC article.
-
A transcriptomic analysis of Echinococcus granulosus larval stages: implications for parasite biology and host adaptation.PLoS Negl Trop Dis. 2012;6(11):e1897. doi: 10.1371/journal.pntd.0001897. Epub 2012 Nov 29. PLoS Negl Trop Dis. 2012. PMID: 23209850 Free PMC article.
-
Comprehensive Expression Profiling of Rice Tetraspanin Genes Reveals Diverse Roles During Development and Abiotic Stress.Front Plant Sci. 2015 Dec 11;6:1088. doi: 10.3389/fpls.2015.01088. eCollection 2015. Front Plant Sci. 2015. PMID: 26697042 Free PMC article.
-
Fine mapping and identification of two NtTOM2A homeologs responsible for tobacco mosaic virus replication in tobacco (Nicotiana tabacum L.).BMC Plant Biol. 2024 Jan 24;24(1):67. doi: 10.1186/s12870-024-04744-y. BMC Plant Biol. 2024. PMID: 38262958 Free PMC article.
References
-
- Boucheix C, Duc GH, Jasmin C, Rubinstein E. Tetraspanins and malignancy. Expert Rev Mol Med. 2001;2001:1–17. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
