

Muscle wasting in aged, sarcopenic rats is associated with enhanced activity of the ubiquitin proteasome pathway
- PMID: 20940294
- PMCID: PMC3000941
- DOI: 10.1074/jbc.M110.129718
Muscle wasting in aged, sarcopenic rats is associated with enhanced activity of the ubiquitin proteasome pathway
Abstract
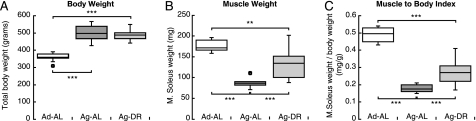
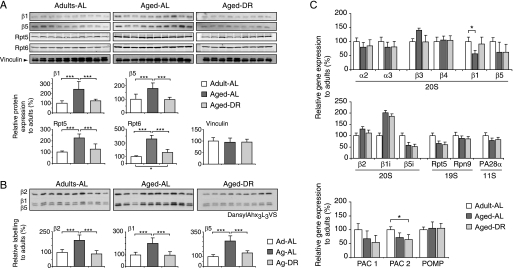
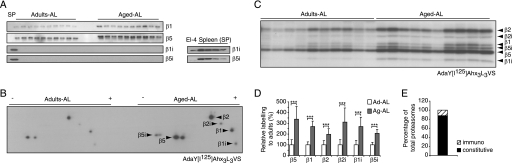
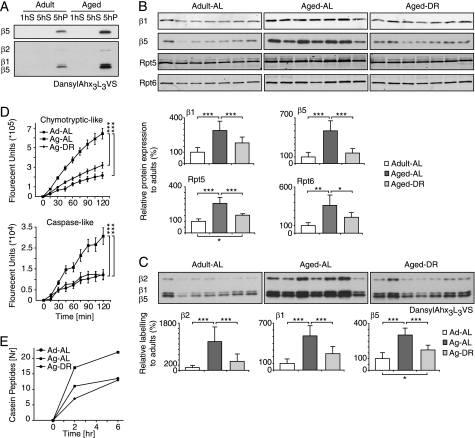
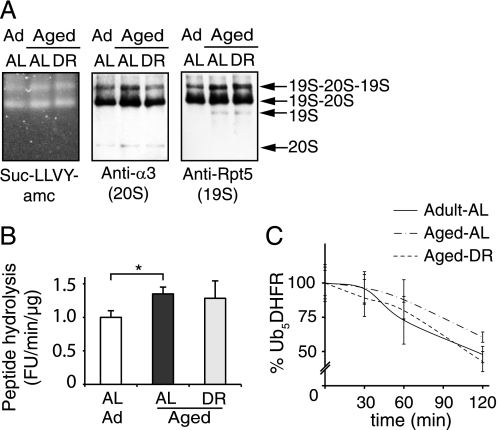
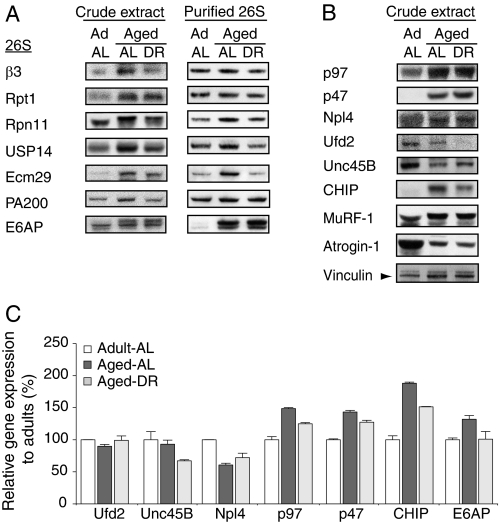
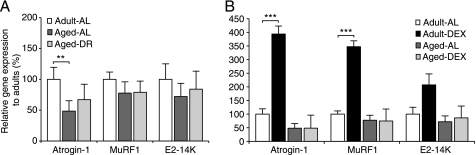
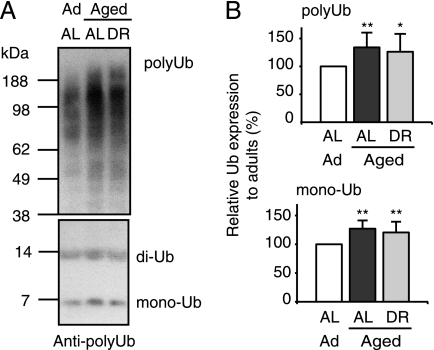
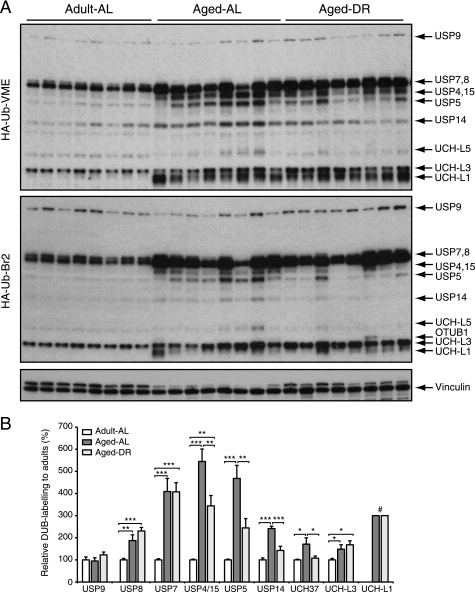
Among the hallmarks of aged organisms are an accumulation of misfolded proteins and a reduction in skeletal muscle mass ("sarcopenia"). We have examined the effects of aging and dietary restriction (which retards many age-related changes) on components of the ubiquitin proteasome system (UPS) in muscle. The hindlimb muscles of aged (30 months old) rats showed a marked loss of muscle mass and contained 2-3-fold higher levels of 26S proteasomes than those of adult (4 months old) controls. 26S proteasomes purified from muscles of aged and adult rats showed a similar capacity to degrade peptides, proteins, and an ubiquitylated substrate, but differed in levels of proteasome-associated proteins (e.g. the ubiquitin ligase E6AP and deubiquitylating enzyme USP14). Also, the activities of many other deubiquitylating enzymes were greatly enhanced in the aged muscles. Nevertheless, their content of polyubiquitylated proteins was higher than in adult animals. The aged muscles contained higher levels of the ubiquitin ligase CHIP, involved in eliminating misfolded proteins, and MuRF1, which ubiquitylates myofibrillar proteins. These muscles differed from ones rapidly atrophying due to disease, fasting, or disuse in that Atrogin-1/MAFbx expression was low and not inducible by glucocorticoids. Thus, the muscles of aged rats showed many adaptations indicating enhanced proteolysis by the UPS, which may enhance their capacity to eliminate misfolded proteins and seems to contribute to the sarcopenia. Accordingly, dietary restriction decreased or prevented the aging-associated increases in proteasomes and other UPS components and reduced muscle wasting.
Figures
References
-
- Evans W. J. (1995) J. Gerontol. A Biol. Sci. Med. Sci. 50, 5–8 - PubMed
-
- Lecker S. H., Goldberg A. L., Mitch W. E. (2006) J. Am. Soc. Nephrol. 17, 1807–1819 - PubMed
-
- Rock K. L., Gramm C., Rothstein L., Clark K., Stein R., Dick L., Hwang D., Goldberg A. L. (1994) Cell 78, 761–771 - PubMed
-
- Voges D., Zwickl P., Baumeister W. (1999) Annu. Rev. Biochem. 68, 1015–1068 - PubMed
-
- Ciechanover A. (2005) Nat. Rev. Mol. Cell Biol. 6, 79–87 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
