

Review
doi: 10.1152/physiol.00002.2010.
Energy landscapes as a tool to integrate GPCR structure, dynamics, and function
Affiliations
- PMID: 20940434
- PMCID: PMC3056154
- DOI: 10.1152/physiol.00002.2010
Item in Clipboard
Review
Energy landscapes as a tool to integrate GPCR structure, dynamics, and function
Physiology (Bethesda).
2010 Oct.
Abstract
G protein-coupled receptors (GPCRs) are versatile signaling molecules that mediate the majority of physiological responses to hormones and neurotransmitters. Recent high-resolution structural insights into GPCR structure and dynamics are beginning to shed light on the molecular basis of this versatility. We use energy landscapes to conceptualize the link between structure and function.
Figures
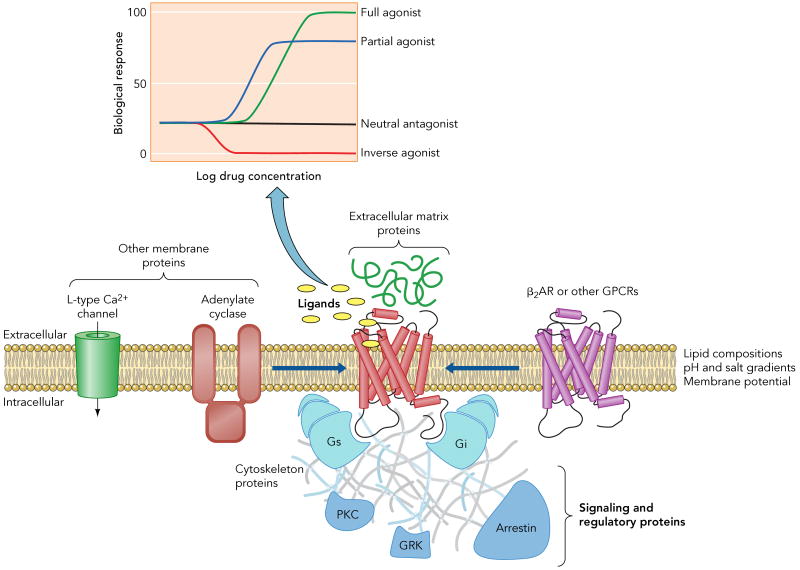
Ligands with different efficacies (inset) can modulate a given signaling pathway. Moreover, the efficacy of a drug may differ for different down stream signaling profiles. For instance, certain drugs induce routes involving signaling and regulatory proteins (e.g. Gi, arrestin) that complement the predominant pathway through Gs. In addition to ligand binding, other factors as lipid bilayer composition, pH and salt gradients, membrane potential, protein-protein interactions, either within the membrane (homo- or hetero-oligomerization and interaction with other membrane proteins) or with extracellular and intracellular partners, or post-translational modifications, as glycosilation, phosphorylation or acylation, can also influence the signaling behavior of the receptor.
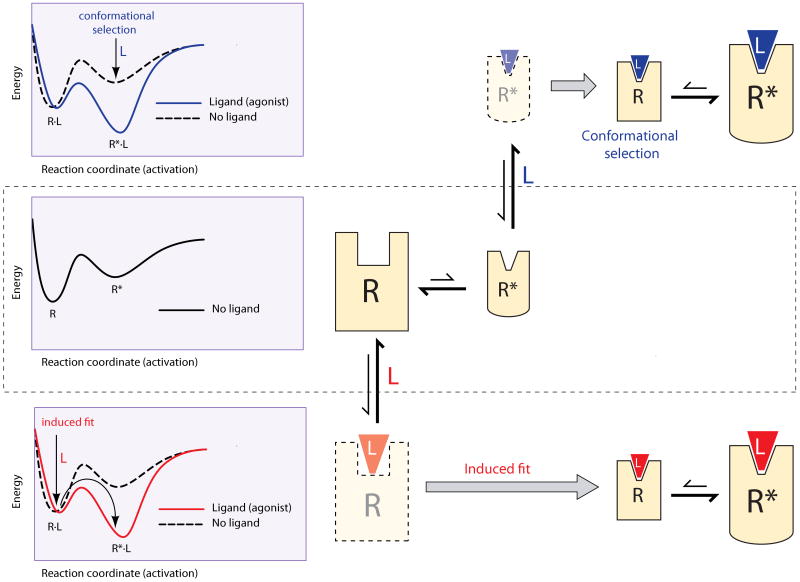
The left panels represent the energy of the receptor along the activation pathway. The minima in the landscapes correspond to stable conformations, connected through saddle points representing conformational changes. The right panels represent the stable conformations in each landscape, where size is proportional to population of the conformer. Translucent figures correspond to transient conformations. The middle panel shows a receptor that in the absence of ligand exists in two conformations, R and R*, with a higher population of the lowest energy conformer, R. In an induced-fit scenario (bottom), the ligand binds to the conformation R to create the R·L complex. Ligand binding alters the energy landscape of the complex (red line), which accelerates activation, stabilizes the complex and provides the energy to induce the conformational changes to R*·L (red arrows). These effects result in an increase of the population of the R*·L conformation. In conformational selection (top panel), the ligand binds to and stabilizes the active conformation R*. Alteration of the energy landscape (blue line) results in a population shift of the receptor conformations towards the R*·L active state.
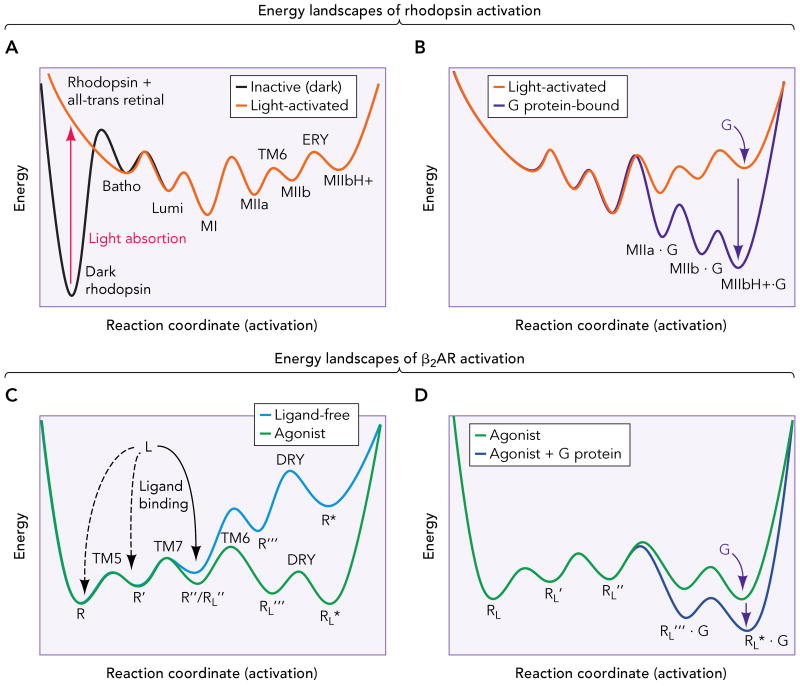
A. Inactive (dark) rhodopsin is trapped in a deep energy well (broken line). Retinal isomerization provides the energy for the receptor to overcome the high initial activation barrier and proceed along the energy landscape through small conformational changes (Batho and Lumi) to the Meta I state (MI). Subsequently, activation proceeds by formation of the Meta IIa (MIIa), Meta IIb (MIIb, by rigid-body movement of TM6) and Meta IIbH+ (MIIbH+, by neutralization of Glu134(3.49) in the ERY motif of TM3) intermediates, and establishment of an equilibrium between the Meta forms. B. G protein binding further changes the energy landscape (black line), displacing the equilibrium to the active ternary complex, capable of catalyze the GDP-GTP exchange in the G protein. C. The β2AR possesses a shallow energy landscape, with several conformational states (R, R′, R″, that differ in small structural changes in TM5 and TM7) separated by relatively low energy barriers (broken line). This translates in an inherent flexibility that allows the ligand-free receptor to explore different conformations. Ligand binding to certain intermediates (R″ in this example) changes the shape of the energy landscape (solid line), and activation proceeds to populate conformations of lower energy (R‴L and R*L). These conformations probably involve a similar set of conformational changes than rhodopsin, i.e. rearrangement of TM6 and neutralization of Asp130(3.49) in the DRY motif of TM3. D. Binding of the G protein to these latter states further changes the energy landscape (black line), lowering the energy and stabilizing the active ternary complex.

This figure represents epinephrine (shown as spheres) bound to the β2AR (TM5, green; TM6, blue; TM7/helix 8, cyan). Receptor activation comprises a series of conformational changes that aggregate into specific activation pathways. Agonists interact with a group of Ser residues on the extracellular side of TM5 (Ser203(5.42), Ser204(5.43) and Ser207(5.46)) involved in ligand binding and receptor activation (44, 45). This interaction disrupts the TM3/TM5 interface at the binding site (35), and these local changes are transmitted to the cytoplasmic side of the helix through the rearrangement of the Pro-induced distortion of TM5 (green arrow) (Sansuk et al., Mol Pharmacol under review)(47). Agonists also induce/stabilize the rigid body movement of TM6 (blue arrow) through their action on Trp(6.48). A complex hydrogen bond network between highly conserved residues and a cluster of water molecules in the TM2-TM6-TM7 interface seems responsible for stabilizing the pi-helix segment observed in the inactive state of rhodopsin(26). This network is most likely conserved in other Class A GPCRs (1), and has a structural and functional role (34). Specifically, we suggest that agonist binding induces changes in this network, which results in local structural changes in the NPxxY motif and in a relocation of the cytoplamic side of TM7 (cyan arrow). These pathways involve highly conserved residues, and are probably conserved in other Family A GPCRs. This figure has been created with PyMOL (DeLano, W.L. The PyMOL Molecular Graphics System (2002) on World Wide Web http://www.pymol.org ).
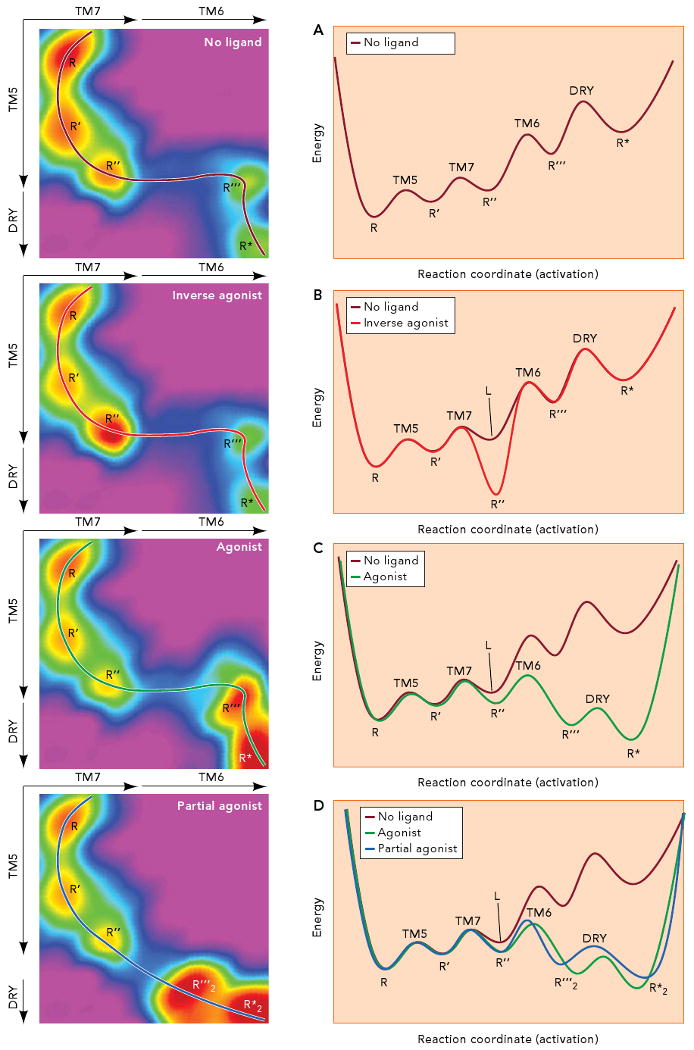
The left panels represent energy in the z axis (color) as a function of the conformational changes in TM7 and TM6 (in the x axis), and of the conformational changes in TM5 and neutralization of Asp130(3.49) in the DRY motif (in the y axis). Activation follows the path traced by the lines in the 3D landscapes. Slicing the 3D surface along these paths results into the simpler 2D landscapes depicted in the right panels. A. In the absence of ligand, small conformational changes in the TM3/TM5 interface and plasticity in the water cluster around the NPxxY motif result in a receptor population that fluctuates between the R, R′ and R″ conformations. The conformations R‴ and R* can get populated by movement of TM6 and neutralization of Asp130(3.49) in the DRY motif of TM3. The R* conformation is able to bind to and activate the G protein, resulting in certain constitutive activity. B. Binding of an inverse agonist stabilizes the R″ conformation, where TM5 and TM7 are slightly rearranged, resulting in a change of the receptor energy landscape (compare panels A and B). As a result, the receptor gets trapped in the R″ conformation. C. An agonist binds to and select the R″ conformation and changes the energy landscape in such a way that the barrier to reach R‴ (i.e. to move TM6) is lowered. The stabilized open crevice in the cytoplasmic side favors the neutralization of Asp130(3.49) in the DRY motif so Arg131(3.50) can reach the extended conformation needed for binding of the G protein. D. A partial agonist binds to and stabilizes R″ to induce a change in the energy landscape. In this case, a conformation R‴2, different from the R‴ conformation stabilized by agonists, becomes available. In this new energy landscape (compare panels C and D), the lower energy conformation corresponds to R*2, which can be somewhat different from R*, but with enough overlapping so it is also able to bind to and activate the G protein. Thus, the stabilization of a new energy landscape implies that the receptor must follow a different activation pathway (compare the black and grey lines in panels C and D).
References
-
- Ballesteros JA, Jensen AD, Liapakis G, Rasmussen SG, Shi L, Gether U, Javitch JA. Activation of the beta 2-adrenergic receptor involves disruption of an ionic lock between the cytoplasmic ends of transmembrane segments 3 and 6. J Biol Chem. 2001;276:29171–29177. - PubMed
-
- Ballesteros JA, Weinstein H. Integrated methods for the construction of three dimensional models and computational probing of structure function relations in G protein-coupled receptors. In: Sealfon SC, Conn PM, editors. Methods in Neurosciences. San Diego, CA: Academic Press; 1995. pp. 366–428.
-
- Bartl FJ, Vogel R. Structural and functional properties of metarhodopsin III: recent spectroscopic studies on deactivation pathways of rhodopsin. Phys Chem Chem Phys. 2007;9:1648–1658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources