

Lungfishes, like tetrapods, possess a vomeronasal system
- PMID: 20941371
- PMCID: PMC2951178
- DOI: 10.3389/fnana.2010.00130
Lungfishes, like tetrapods, possess a vomeronasal system
Abstract
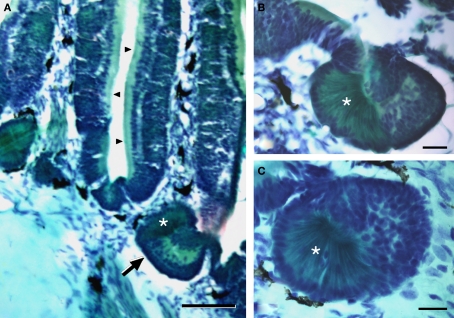
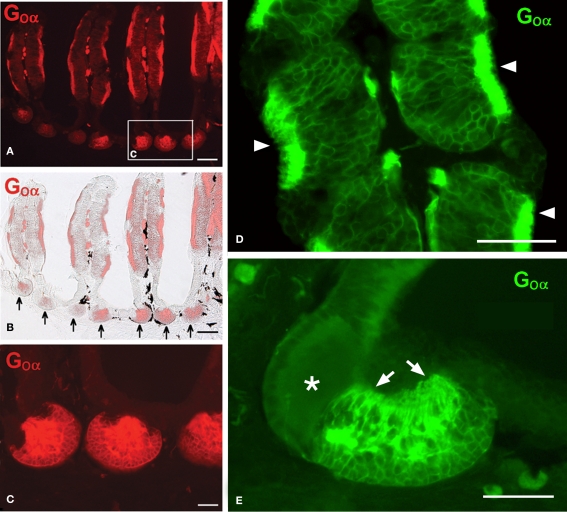
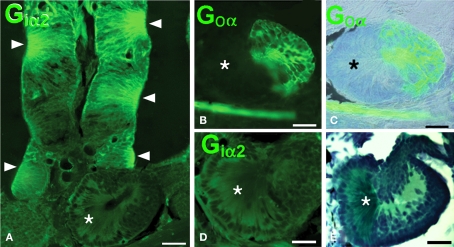
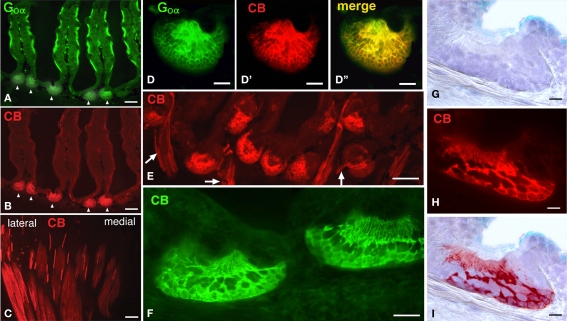
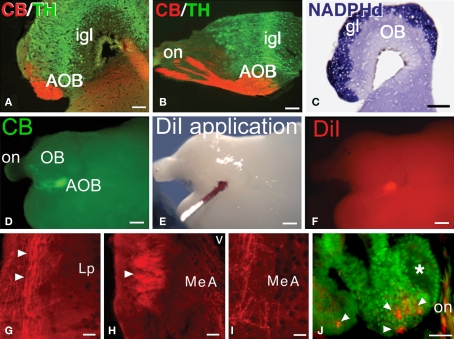
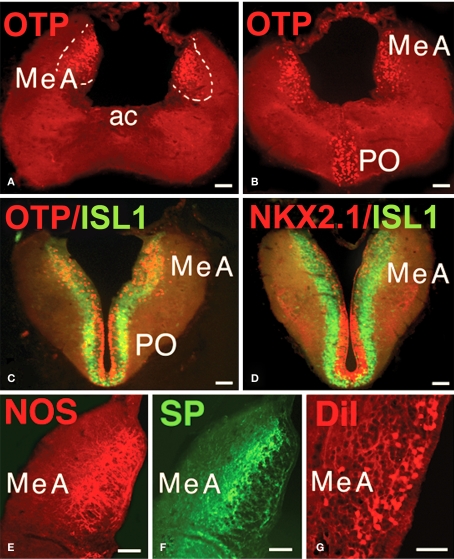
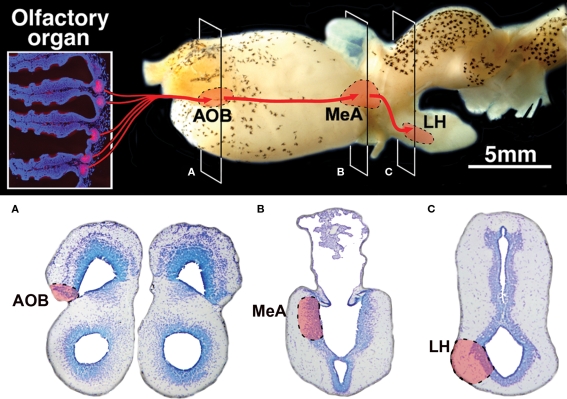
The vomeronasal system (VNS) is an accessory olfactory system that in tetrapod vertebrates is composed of specific receptor neurons in the nasal organ and a set of centers in the forebrain that receive and relay the information consecutively towards the hypothalamus. Thus, only in tetrapods the VNS comprises a discrete vomeronasal (Jacobson's) organ, which contains receptor cells that are morphologically distinct from those of the olfactory epithelium and use different transduction mechanisms. The axons of the vomeronasal receptors in tetrapods project to the accessory olfactory bulb (AOB) in the rostral telencephalon. Secondary vomeronasal connections exist through the medial amygdala to the hypothalamus. Currently, the lungfishes are considered the closest living relatives of tetrapods. Here we show that the African lungfish, Protopterus dolloi, has epithelial crypts at the base of the lamellae of the olfactory epithelium that express markers of the vomeronasal receptors in tetrapods. The projections of these crypts allow us to identify an AOB on the lateral margin of the main olfactory bulb. The projections of this AOB reach a region that is topologically, hodologically, and immunohistochemically identical to the medial amygdala and could represent its homolog. Neurons of this putative medial amygdala were demonstrated to project to the lateral hypothalamus, as they do in tetrapods. All these features that lungfishes share with tetrapods indicate that lungfishes have the complete set of brain centers and connections involved in processing vomeronasal information and that these features were already present in the last common ancestor of lungfishes and tetrapods.
Keywords: accessory olfactory bulb; evolution; homology; lobe-finned fish; medial amygdala.
Figures
Similar articles
-
Histological and ultrastructural characteristics of the primordial vomeronasal organ in lungfish.Anat Rec (Hoboken). 2012 Mar;295(3):481-91. doi: 10.1002/ar.22415. Epub 2012 Jan 23. Anat Rec (Hoboken). 2012. PMID: 22271496
-
Sex pheromone systems in goldfish: comparisons to vomeronasal systems in tetrapods.Brain Behav Evol. 1993;42(4-5):265-80. doi: 10.1159/000114166. Brain Behav Evol. 1993. PMID: 8252378 Review.
-
Lateral and medial amygdala of anuran amphibians and their relation to olfactory and vomeronasal information.Brain Res Bull. 2005 Sep 15;66(4-6):332-6. doi: 10.1016/j.brainresbull.2005.05.017. Brain Res Bull. 2005. PMID: 16144609
-
Olfactory projections in the lepidosirenid lungfishes.Brain Behav Evol. 2012;79(1):4-25. doi: 10.1159/000331267. Epub 2011 Nov 8. Brain Behav Evol. 2012. PMID: 22067508
-
Molecules, fossils, and the origin of tetrapods.J Mol Evol. 1992 Aug;35(2):102-13. doi: 10.1007/BF00183221. J Mol Evol. 1992. PMID: 1501250 Review.
Cited by
-
Distribution of recesses in the olfactory organ of African lungfish Protopterus aethiopicus.J Vet Med Sci. 2022 Jul 1;84(7):885-889. doi: 10.1292/jvms.22-0173. Epub 2022 May 5. J Vet Med Sci. 2022. PMID: 35527019 Free PMC article.
-
Expression of SATB1 and SATB2 in the brain of bony fishes: what fish reveal about evolution.Brain Struct Funct. 2023 May;228(3-4):921-945. doi: 10.1007/s00429-023-02632-z. Epub 2023 Apr 1. Brain Struct Funct. 2023. PMID: 37002478 Free PMC article.
-
Mitral cell development in the olfactory bulb of sharks: evidences of a conserved pattern of glutamatergic neurogenesis.Brain Struct Funct. 2019 Sep;224(7):2325-2341. doi: 10.1007/s00429-019-01906-9. Epub 2019 Jun 15. Brain Struct Funct. 2019. PMID: 31203451 Free PMC article.
-
A detailed 3D MRI brain atlas of the African lungfish Protopterus annectens.Sci Rep. 2024 Apr 5;14(1):7999. doi: 10.1038/s41598-024-58671-x. Sci Rep. 2024. PMID: 38580713 Free PMC article.
-
The Zebrafish Amygdaloid Complex - Functional Ground Plan, Molecular Delineation, and Everted Topology.Front Neurosci. 2020 Jul 16;14:608. doi: 10.3389/fnins.2020.00608. eCollection 2020. Front Neurosci. 2020. PMID: 32765204 Free PMC article.
References
LinkOut - more resources
Full Text Sources