

Sequestration and tissue accumulation of human malaria parasites: can we learn anything from rodent models of malaria?
- PMID: 20941396
- PMCID: PMC2947991
- DOI: 10.1371/journal.ppat.1001032
Sequestration and tissue accumulation of human malaria parasites: can we learn anything from rodent models of malaria?
Abstract
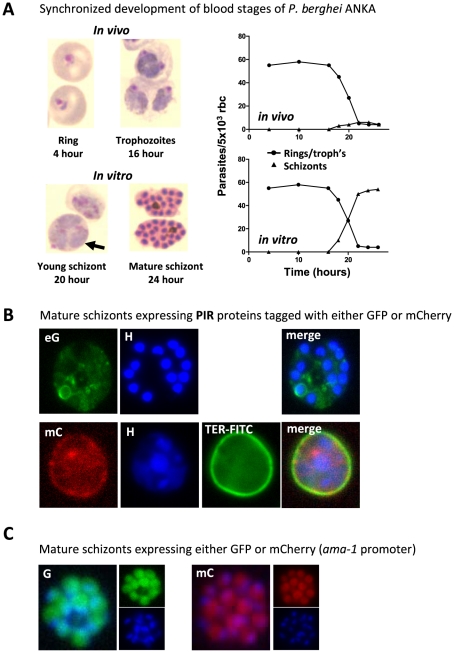
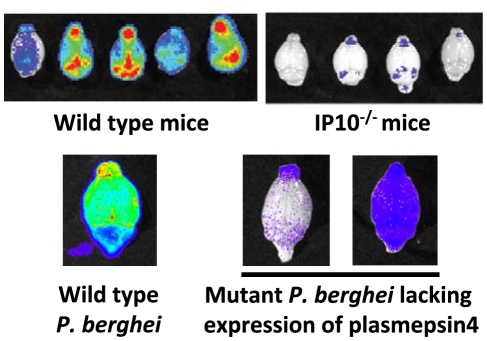
The sequestration of Plasmodium falciparum-infected red blood cells (irbcs) in the microvasculature of organs is associated with severe disease; correspondingly, the molecular basis of irbc adherence is an active area of study. In contrast to P. falciparum, much less is known about sequestration in other Plasmodium parasites, including those species that are used as models to study severe malaria. Here, we review the cytoadherence properties of irbcs of the rodent parasite Plasmodium berghei ANKA, where schizonts demonstrate a clear sequestration phenotype. Real-time in vivo imaging of transgenic P. berghei parasites in rodents has revealed a CD36-dependent sequestration in lungs and adipose tissue. In the absence of direct orthologs of the P. falciparum proteins that mediate binding to human CD36, the P. berghei proteins and/or mechanisms of rodent CD36 binding are as yet unknown. In addition to CD36-dependent schizont sequestration, irbcs accumulate during severe disease in different tissues, including the brain. The role of sequestration is discussed in the context of disease as are the general (dis)similarities of P. berghei and P. falciparum sequestration.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Rogerson SJ, Hviid L, Duffy PE, Leke RF, Taylor DW. Malaria in pregnancy: pathogenesis and immunity. Lancet Infect Dis. 2007;7:105–117. - PubMed
-
- Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, et al. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis. 2007;7:93–104. - PubMed
-
- Beeson JG, Duffy PE. The immunology and pathogenesis of malaria during pregnancy. Curr Top Microbiol Immunol. 2005;297:187–227. - PubMed
-
- Mackintosh CL, Beeson JG, Marsh K. Clinical features and pathogenesis of severe malaria. Trends Parasitol. 2004;20:597–603. - PubMed
-
- Rasti N, Wahlgren M, Chen Q. Molecular aspects of malaria pathogenesis. FEMS Immunol Med Microbiol. 2004;41:9–26. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
