

Dual function of MyD88 in RAS signaling and inflammation, leading to mouse and human cell transformation
- PMID: 20941850
- PMCID: PMC2947228
- DOI: 10.1172/jci42771
Dual function of MyD88 in RAS signaling and inflammation, leading to mouse and human cell transformation
Abstract
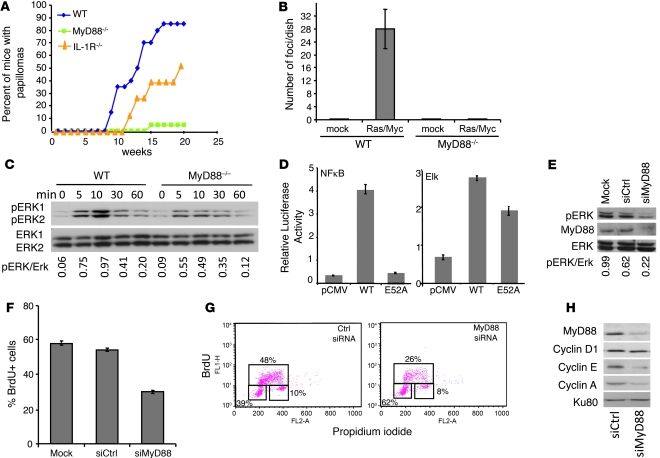
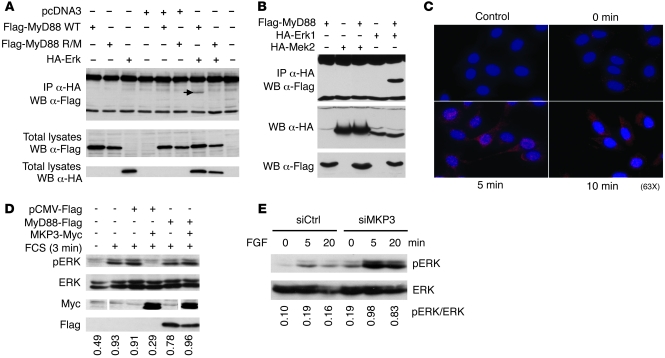
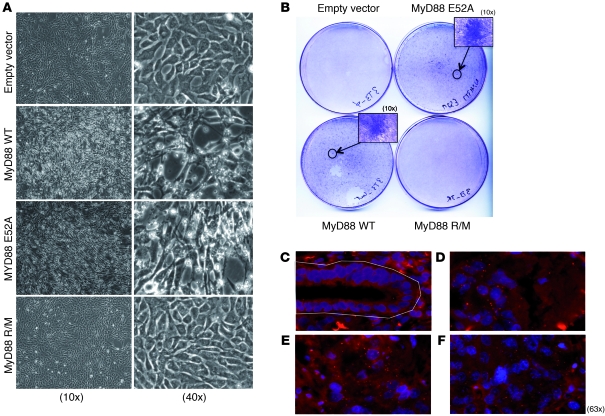
Accumulating evidence points to inflammation as a promoter of carcinogenesis. MyD88 is an adaptor molecule in TLR and IL-1R signaling that was recently implicated in tumorigenesis through proinflammatory mechanisms. Here we have shown that MyD88 is also required in a cell-autonomous fashion for RAS-mediated carcinogenesis in mice in vivo and for MAPK activation and transformation in vitro. Mechanistically, MyD88 bound to the key MAPK, Erk, and prevented its inactivation by its phosphatase, MKP3, thereby amplifying the activation of the canonical RAS pathway. The relevance of this mechanism to human neoplasia was suggested by the finding that MyD88 was overexpressed and interacted with activated Erk in primary human cancer tissues. Collectively, these results show that in addition to its role in inflammation, MyD88 plays what we believe to be a crucial direct role in RAS signaling, cell-cycle control, and cell transformation.
Figures
References
-
- Rakoff-Nahoum S, Medzhitov R. Regulation of spontaneous intestinal tumorigenesis through the adaptor protein MyD88. Science. 2007;317(5834):124–127. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
