

Lipid signaling in T-cell development and function
- PMID: 20943760
- PMCID: PMC2964181
- DOI: 10.1101/cshperspect.a002428
Lipid signaling in T-cell development and function
Abstract
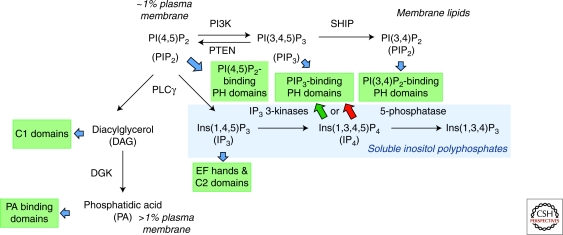
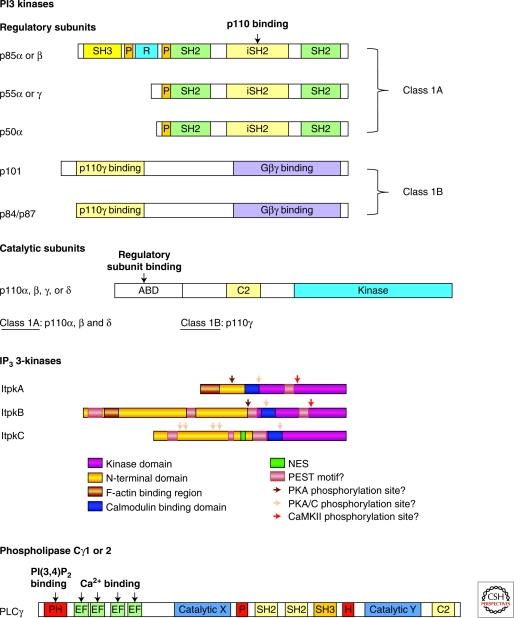
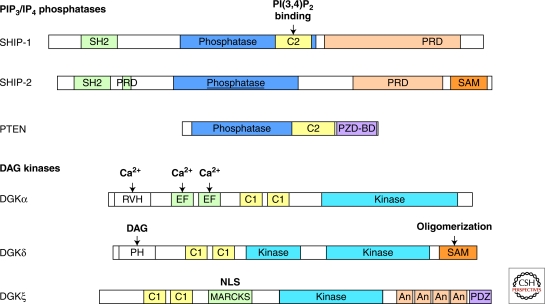
Second messenger molecules relay, amplify, and diversify cell surface receptor signals. Two important examples are phosphorylated D-myo-inositol derivatives, such as phosphoinositide lipids within cellular membranes, and soluble inositol phosphates. Here, we review how phosphoinositide metabolism generates multiple second messengers with important roles in T-cell development and function. They include soluble inositol(1,4,5)trisphosphate, long known for its Ca(2+)-mobilizing function, and phosphatidylinositol(3,4,5)trisphosphate, whose generation by phosphoinositide 3-kinase and turnover by the phosphatases PTEN and SHIP control a key "hub" of TCR signaling. More recent studies unveiled important second messenger functions for diacylglycerol, phosphatidic acid, and soluble inositol(1,3,4,5)tetrakisphosphate (IP(4)) in immune cells. Inositol(1,3,4,5)tetrakisphosphate acts as a soluble phosphatidylinositol(3,4,5)trisphosphate analog to control protein membrane recruitment. We propose that phosphoinositide lipids and soluble inositol phosphates (IPs) can act as complementary partners whose interplay could have broadly important roles in cellular signaling.
Figures
References
-
- Alcazar-Roman AR, Wente SR 2008. Inositol polyphosphates: a new frontier for regulating gene expression. Chromosoma 117: 1–13 - PubMed
-
- Atherly LO, Lucas JA, Felices M, Yin CC, Reiner SL, Berg LJ 2006. The Tec family tyrosine kinases Itk and Rlk regulate the development of conventional CD8+ T cells. Immunity 25: 79–91 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous