

Prototypic seizure activity driven by mature hippocampal fast-spiking interneurons
- PMID: 20943908
- PMCID: PMC6633708
- DOI: 10.1523/JNEUROSCI.1523-10.2010
Prototypic seizure activity driven by mature hippocampal fast-spiking interneurons
Abstract
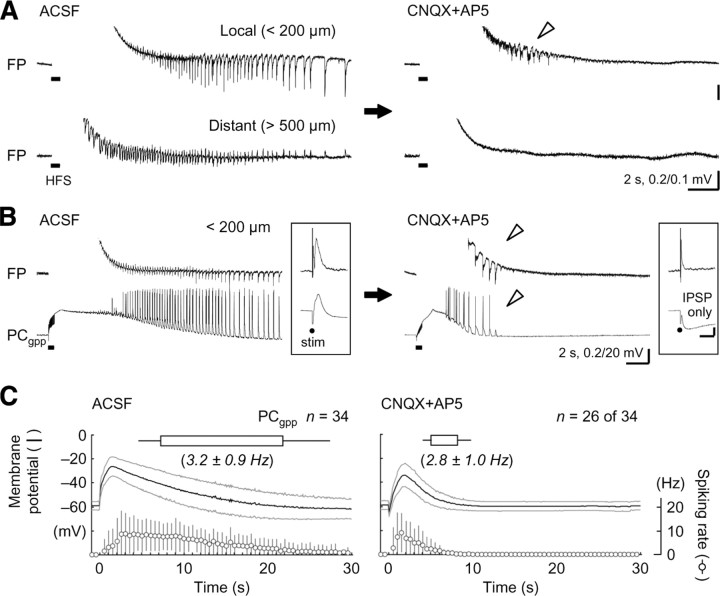
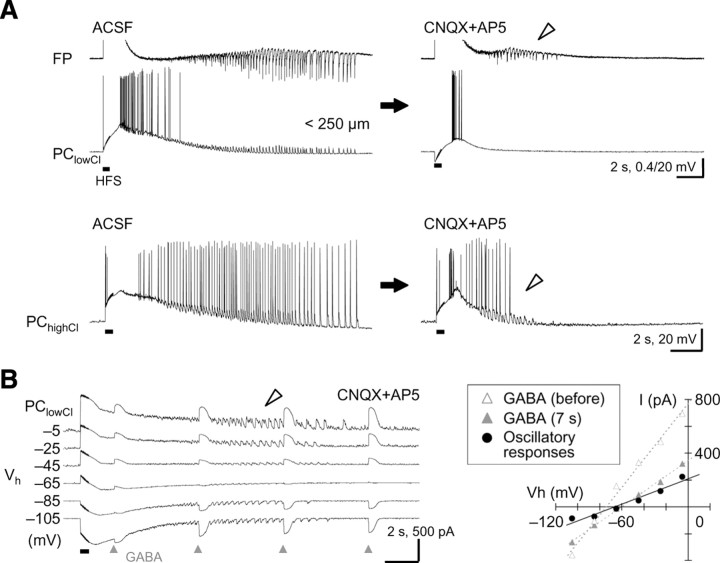
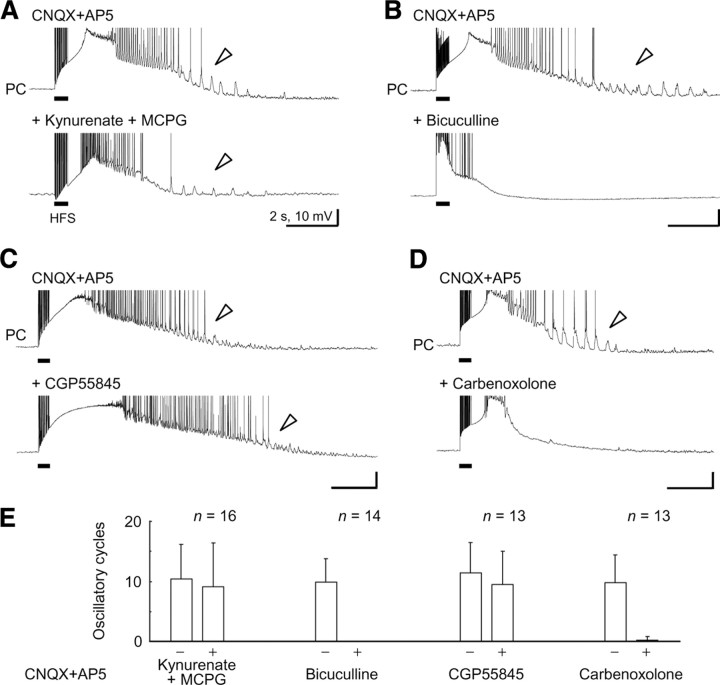
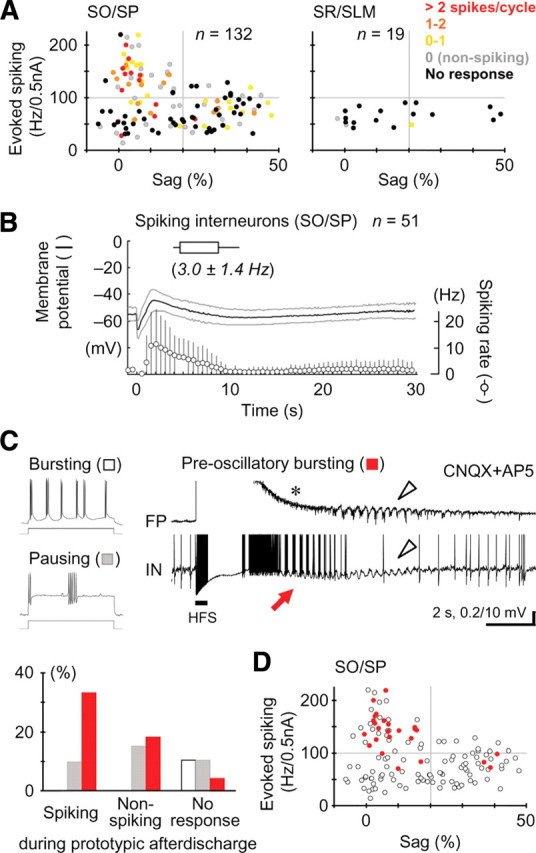
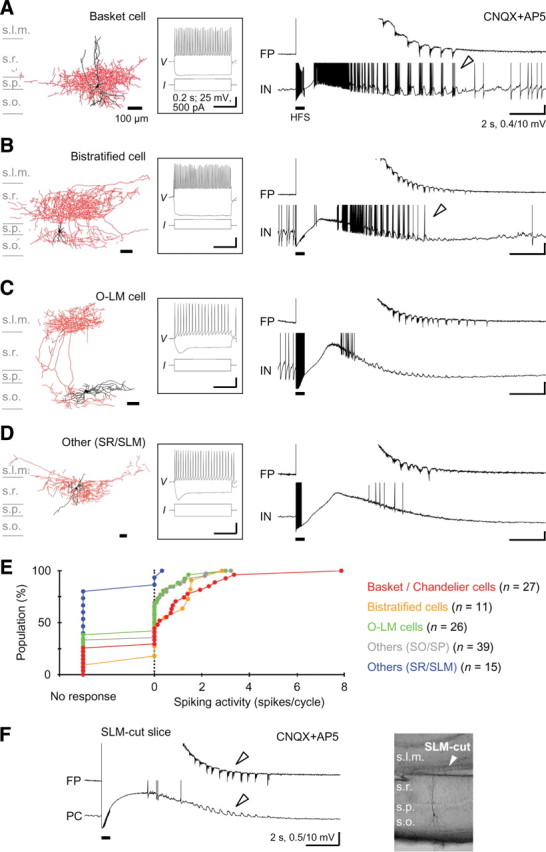
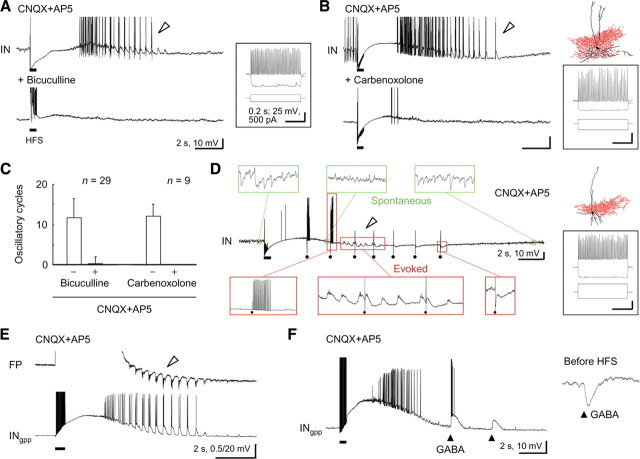
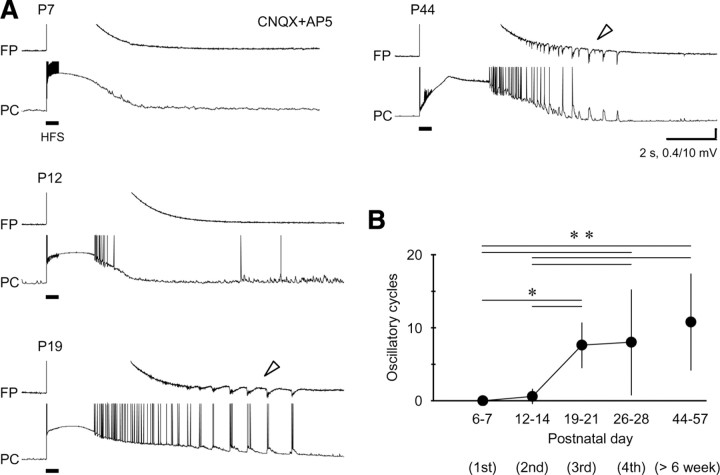
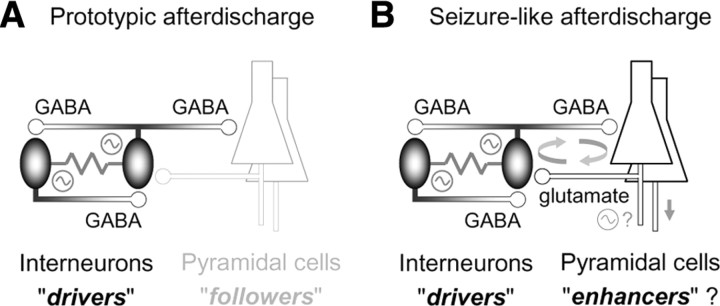
A variety of epileptic seizure models have shown that activation of glutamatergic pyramidal cells is usually required for rhythm generation and/or synchronization in hippocampal seizure-like oscillations in vitro. However, it still remains unclear whether GABAergic interneurons may be able to drive the seizure-like oscillations without glutamatergic transmission. Here, we found that electrical stimulation in rat hippocampal CA1 slices induced a putative prototype of seizure-like oscillations ("prototypic afterdischarge," 1.8-3.8 Hz) in mature pyramidal cells and interneurons in the presence of ionotropic glutamate receptor antagonists. The prototypic afterdischarge was abolished by GABA(A) receptor antagonists or gap junction blockers, but not by a metabotropic glutamate receptor antagonist or a GABA(B) receptor antagonist. Gramicidin-perforated patch-clamp and voltage-clamp recordings revealed that pyramidal cells were depolarized and frequently excited directly through excitatory GABAergic transmissions in each cycle of the prototypic afterdischarge. Interneurons that were actively spiking during the prototypic afterdischarge were mostly fast-spiking (FS) interneurons located in the strata oriens and pyramidale. Morphologically, these interneurons that might be "potential seizure drivers" included basket, chandelier, and bistratified cells. Furthermore, they received direct excitatory GABAergic input during the prototypic afterdischarge. The O-LM cells and most of the interneurons in the strata radiatum and lacunosum moleculare were not essential for the generation of prototypic afterdischarge. The GABA-mediated prototypic afterdischarge was observed later than the third postnatal week in the rat hippocampus. Our results suggest that an FS interneuron network alone can drive the prototypic form of electrically induced seizure-like oscillations through their excitatory GABAergic transmissions and presumably through gap junction-mediated communications.
Figures
Similar articles
-
Synaptic interactions between pyramidal cells and interneurone subtypes during seizure-like activity in the rat hippocampus.J Physiol. 2004 Jun 15;557(Pt 3):961-79. doi: 10.1113/jphysiol.2003.059915. Epub 2004 Apr 23. J Physiol. 2004. PMID: 15107470 Free PMC article.
-
[Neural mechanism underlying generation of synchronous oscillations in hippocampal network].Brain Nerve. 2008 Jul;60(7):755-62. Brain Nerve. 2008. PMID: 18646615 Review. Japanese.
-
Excitatory GABA input directly drives seizure-like rhythmic synchronization in mature hippocampal CA1 pyramidal cells.Neuroscience. 2003;119(1):265-75. doi: 10.1016/s0306-4522(03)00102-7. Neuroscience. 2003. PMID: 12763087
-
Use-dependent shift from inhibitory to excitatory GABAA receptor action in SP-O interneurons in the rat hippocampal CA3 area.J Neurophysiol. 2003 Sep;90(3):1983-95. doi: 10.1152/jn.00060.2003. Epub 2003 May 15. J Neurophysiol. 2003. PMID: 12750426
-
A network mechanism underlying hippocampal seizure-like synchronous oscillations.Neurosci Res. 2008 Jul;61(3):227-33. doi: 10.1016/j.neures.2008.04.002. Epub 2008 Apr 20. Neurosci Res. 2008. PMID: 18457889 Review.
Cited by
-
Divergent paths to seizure-like events.Physiol Rep. 2019 Oct;7(19):e14226. doi: 10.14814/phy2.14226. Physiol Rep. 2019. PMID: 31587522 Free PMC article.
-
Learning to define an electrical biomarker of the epileptogenic zone.Hum Brain Mapp. 2020 Feb 1;41(2):429-441. doi: 10.1002/hbm.24813. Epub 2019 Oct 14. Hum Brain Mapp. 2020. PMID: 31609058 Free PMC article.
-
Optogenetic Approaches for Controlling Seizure Activity.Brain Stimul. 2016 Nov-Dec;9(6):801-810. doi: 10.1016/j.brs.2016.06.055. Epub 2016 Jul 14. Brain Stimul. 2016. PMID: 27496002 Free PMC article. Review.
-
Three-dimensional reconstruction of electron micrographs reveals intrabulbar circuit differences between accessory and main olfactory bulbs.Front Neuroanat. 2013 Apr 22;7:5. doi: 10.3389/fnana.2013.00005. eCollection 2013. Front Neuroanat. 2013. PMID: 23626525 Free PMC article.
-
Cell-attached recordings of responses evoked by photorelease of GABA in the immature cortical neurons.Front Cell Neurosci. 2013 May 31;7:83. doi: 10.3389/fncel.2013.00083. eCollection 2013. Front Cell Neurosci. 2013. PMID: 23754981 Free PMC article.
References
-
- Anderson WW, Lewis DV, Swartzwelder HS, Wilson WA. Magnesium-free medium activates seizure-like events in the rat hippocampal slice. Brain Res. 1986;398:215–219. - PubMed
-
- Avoli M, Psarropoulou C, Tancredi V, Fueta Y. On the synchronous activity induced by 4-aminopyridine in the CA3 subfield of juvenile rat hippocampus. J Neurophysiol. 1993;70:1018–1029. - PubMed
-
- Ben-Ari Y. Excitatory actions of GABA during development: the nature of the nurture. Nat Rev Neurosci. 2002;3:728–739. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous