

Fitness disadvantage of transitional intermediates contributes to dynamic change in the infecting-virus population during coreceptor switch in R5 simian/human immunodeficiency virus-infected macaques
- PMID: 20943985
- PMCID: PMC3004298
- DOI: 10.1128/JVI.01478-10
Fitness disadvantage of transitional intermediates contributes to dynamic change in the infecting-virus population during coreceptor switch in R5 simian/human immunodeficiency virus-infected macaques
Abstract
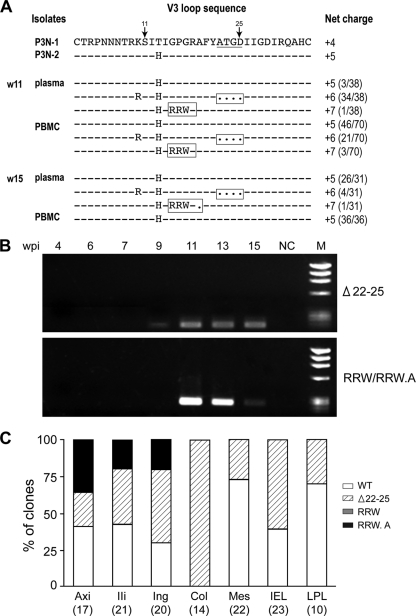
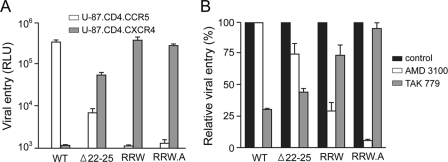
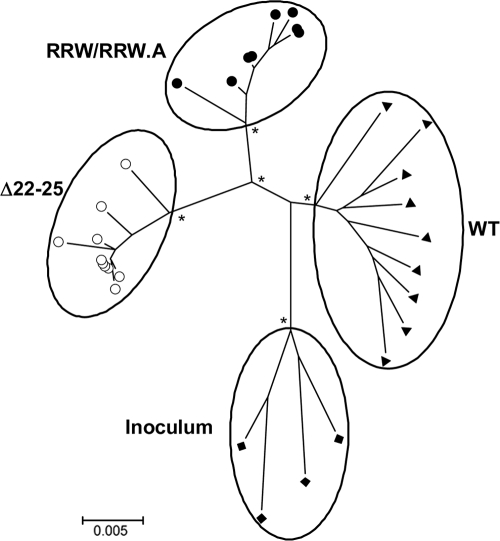
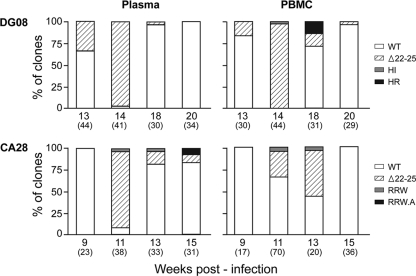
Fitness disadvantage of the transitional intermediates compared to the initial R5 viruses has been suggested to constitute one of the blockades to coreceptor switching, explaining the late appearance of X4 viruses. Using a simian model for human immunodeficiency virus type 1 (HIV-1) coreceptor switching, we demonstrate in this study that similar molecular evolutionary pathways to coreceptor switch occur in more than one R5 simian/human immunodeficiency virus (SHIV)(SF162P3N)-infected macaque. In infected animals where multiple pathways for expansion or switch to CXCR4 coexist, fitness of the transitional intermediates in coreceptor usage efficiency influences their outgrowth and representation in the infecting virus population. Dualtropic and X4 viruses appear at different disease stages, but they have lower entry efficiency than the coexisting R5 strains, which may explain why they do not outcompete the R5 viruses. Similar observations were made in two infected macaques with coreceptor switch, providing in vivo evidence that fitness disadvantage is an obstacle to X4 emergence and expansion.
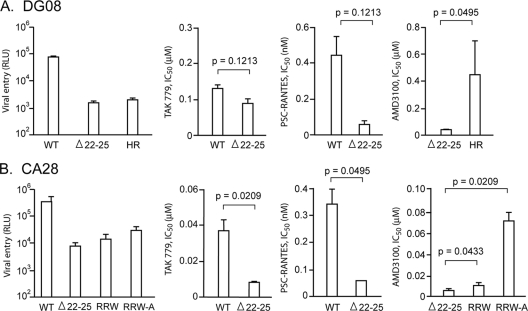
Figures
Similar articles
-
R5X4 viruses are evolutionary, functional, and antigenic intermediates in the pathway of a simian-human immunodeficiency virus coreceptor switch.J Virol. 2008 Jul;82(14):7089-99. doi: 10.1128/JVI.00570-08. Epub 2008 May 14. J Virol. 2008. PMID: 18480460 Free PMC article.
-
Coreceptor switch in R5-tropic simian/human immunodeficiency virus-infected macaques.J Virol. 2007 Aug;81(16):8621-33. doi: 10.1128/JVI.00759-07. Epub 2007 May 30. J Virol. 2007. PMID: 17537860 Free PMC article.
-
V3 loop-determined coreceptor preference dictates the dynamics of CD4+-T-cell loss in simian-human immunodeficiency virus-infected macaques.J Virol. 2005 Oct;79(19):12296-303. doi: 10.1128/JVI.79.19.12296-12303.2005. J Virol. 2005. PMID: 16160156 Free PMC article.
-
Coreceptor switch in infection of nonhuman primates.Curr HIV Res. 2009 Jan;7(1):30-8. doi: 10.2174/157016209787048500. Curr HIV Res. 2009. PMID: 19149552 Review.
-
Genotypic coreceptor analysis.Eur J Med Res. 2007 Oct 15;12(9):453-62. Eur J Med Res. 2007. PMID: 17933727 Review.
Cited by
-
Identification of interdependent variables that influence coreceptor switch in R5 SHIV(SF162P3N)-infected macaques.Retrovirology. 2012 Dec 13;9:106. doi: 10.1186/1742-4690-9-106. Retrovirology. 2012. PMID: 23237529 Free PMC article.
-
Generation of lineage-related, mucosally transmissible subtype C R5 simian-human immunodeficiency viruses capable of AIDS development, induction of neurological disease, and coreceptor switching in rhesus macaques.J Virol. 2013 Jun;87(11):6137-49. doi: 10.1128/JVI.00178-13. Epub 2013 Mar 20. J Virol. 2013. PMID: 23514895 Free PMC article.
-
Stochastic model of in-vivo X4 emergence during HIV infection: implications for the CCR5 inhibitor maraviroc.PLoS One. 2012;7(7):e38755. doi: 10.1371/journal.pone.0038755. Epub 2012 Jul 17. PLoS One. 2012. PMID: 22866173 Free PMC article.
-
Adoption of an "open" envelope conformation facilitating CD4 binding and structural remodeling precedes coreceptor switch in R5 SHIV-infected macaques.PLoS One. 2011;6(7):e21350. doi: 10.1371/journal.pone.0021350. Epub 2011 Jul 8. PLoS One. 2011. PMID: 21760891 Free PMC article.
-
Emergence of CD4 independence envelopes and astrocyte infection in R5 simian-human immunodeficiency virus model of encephalitis.J Virol. 2014 Aug;88(15):8407-20. doi: 10.1128/JVI.01237-14. Epub 2014 May 14. J Virol. 2014. PMID: 24829360 Free PMC article.
References
-
- Agrawal-Gamse, C., F. H. Lee, B. Haggarty, A. P. Jordan, Y. Yi, B. Lee, R. G. Collman, J. A. Hoxie, R. W. Doms, and M. M. Laakso. 2009. Adaptive mutations in a human immunodeficiency virus type 1 envelope protein with a truncated V3 loop restore function by improving interactions with CD4. J. Virol. 83:11005-11015. - PMC - PubMed
-
- Allen, T. M., M. Altfeld, S. C. Geer, E. T. Kalife, C. Moore, M. O'Sullivan, K. I. Desouza, M. E. Feeney, R. L. Eldridge, E. L. Maier, D. E. Kaufmann, M. P. Lahaie, L. Reyor, G. Tanzi, M. N. Johnston, C. Brander, R. Draenert, J. K. Rockstroh, H. Jessen, E. S. Rosenberg, S. A. Mallal, and B. D. Walker. 2005. Selective escape from CD8+ T-cell responses represents a major driving force of human immunodeficiency virus type 1 (HIV-1) sequence diversity and reveals constraints on HIV-1 evolution. J. Virol. 79:13239-13249. - PMC - PubMed
-
- Asjo, B., L. Morfeldt-Manson, J. Albert, G. Biberfeld, A. Karlsson, K. Lidman, and E. M. Fenyo. 1986. Replicative capacity of human immunodeficiency virus from patients with varying severity of HIV infection. Lancet ii:660-662. - PubMed
-
- Berger, E. A., P. M. Murphy, and J. M. Farber. 1999. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu. Rev. Immunol. 17:657-700. - PubMed
-
- Brenchley, J. M., T. W. Schacker, L. E. Ruff, D. A. Price, J. H. Taylor, G. J. Beilman, P. L. Nguyen, A. Khoruts, M. Larson, A. T. Haase, and D. C. Douek. 2004. CD4+ T cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J. Exp. Med. 200:749-759. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
