

Parental division of labor, coordination, and the effects of family structure on parenting in monogamous prairie voles (Microtus ochrogaster)
- PMID: 20945408
- PMCID: PMC3164816
- DOI: 10.1002/dev.20498
Parental division of labor, coordination, and the effects of family structure on parenting in monogamous prairie voles (Microtus ochrogaster)
Abstract
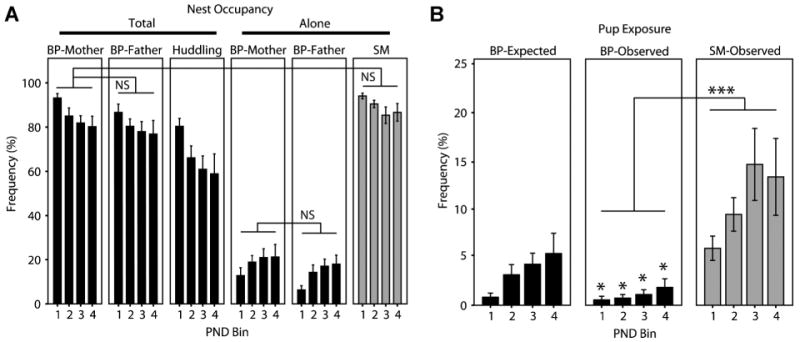
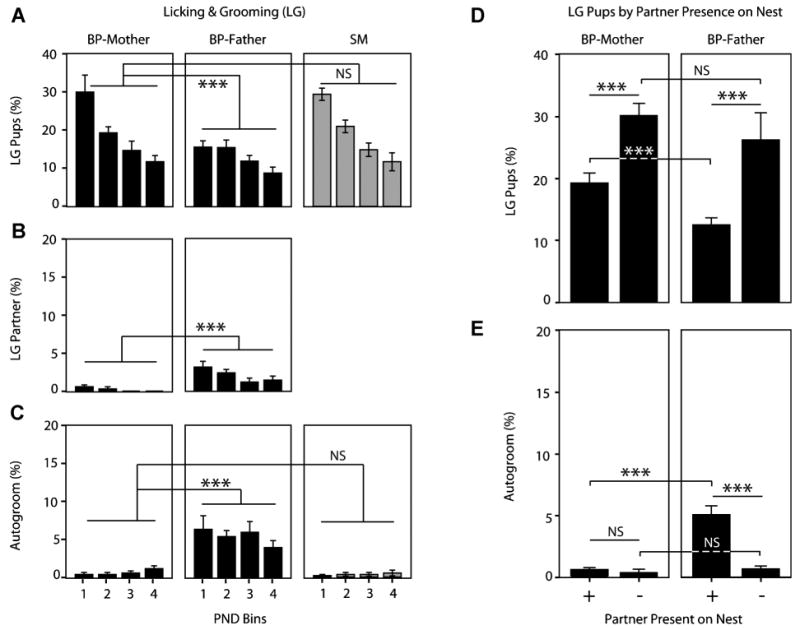
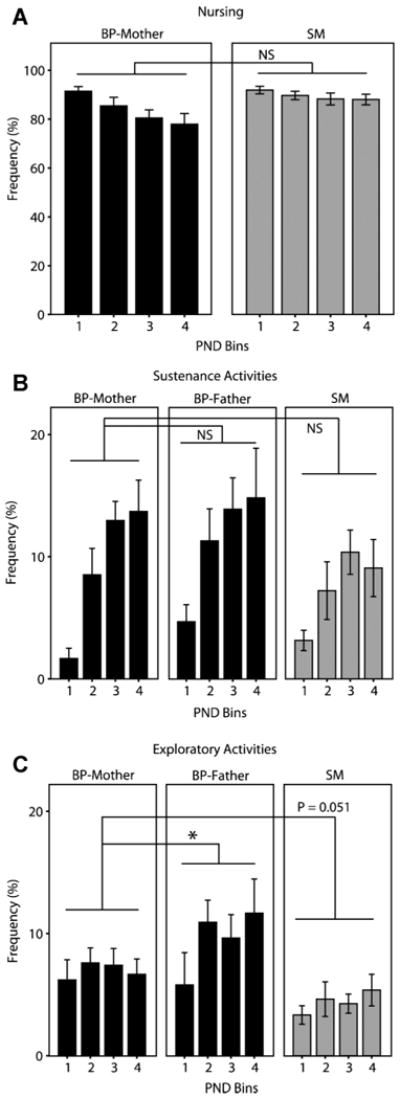
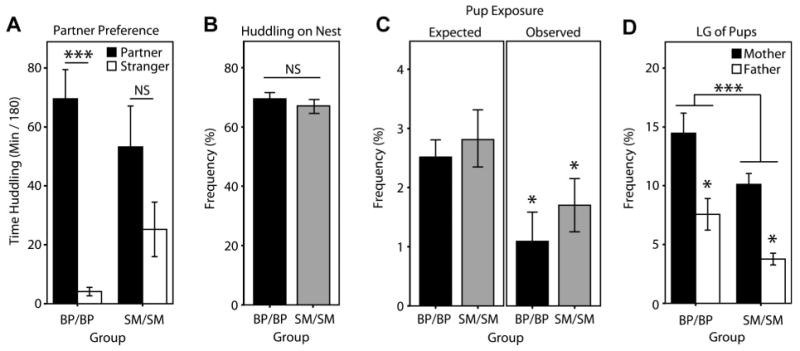
Family relationships help shape species-typical social and emotional development, but our understanding of how this shaping occurs is still relatively limited. Prairie voles are a socially monogamous and biparental species that is well situated to complement traditional animal models, such as rats and mice, in investigating the effects of family experience. In this series of studies, we aimed to test hypotheses relating to how prairie vole families function under undisturbed, standard laboratory conditions. In the first study, we compared the parental behavior of primiparous biparental (BP) and single-mother (SM) prairie vole family units for 12 postnatal days and then tested for sex differences, behavioral coordination, and family structure effects. Under BP conditions, nest attendance was coordinated and shared equally by both sexes, while pup-directed and partner-directed licking and grooming (LG) were coordinated in a sex and social-context-dependent manner. Contrary to our expectations, SMs showed no evidence of strong parental compensation in response to the lack of the father, indicating a minimal effect of family structure on maternal behavior but a large effect on pup care. In the second study, we examined the effects of these BP and SM rearing conditions on family dynamics in the next generation and found that SM-reared adult parents exhibited lower rates of pup-directed LG in comparison to BP-reared counterparts. Situated in the context of human family dynamics and psychology, these results suggest that the study in prairie voles may help improve our understanding of family systems and how perturbations to these systems can affect adults and offspring.
Copyright © 2010 Wiley Periodicals, Inc.
Figures
References
-
- Amato PR. The consequences of divorce for adults and children. Journal of Marriage and Family. 2000;62(4):1269–1287.
-
- Aragona BJ, Liu Y, Yu YJ, Curtis JT, Detwiler JM, Insel TR. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nature Neuroscience. 2006;9(1):133. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
