

Transcellular and paracellular pathways of transepithelial fluid secretion in Malpighian (renal) tubules of the yellow fever mosquito Aedes aegypti
- PMID: 20946239
- PMCID: PMC3032036
- DOI: 10.1111/j.1748-1716.2010.02195.x
Transcellular and paracellular pathways of transepithelial fluid secretion in Malpighian (renal) tubules of the yellow fever mosquito Aedes aegypti
Abstract
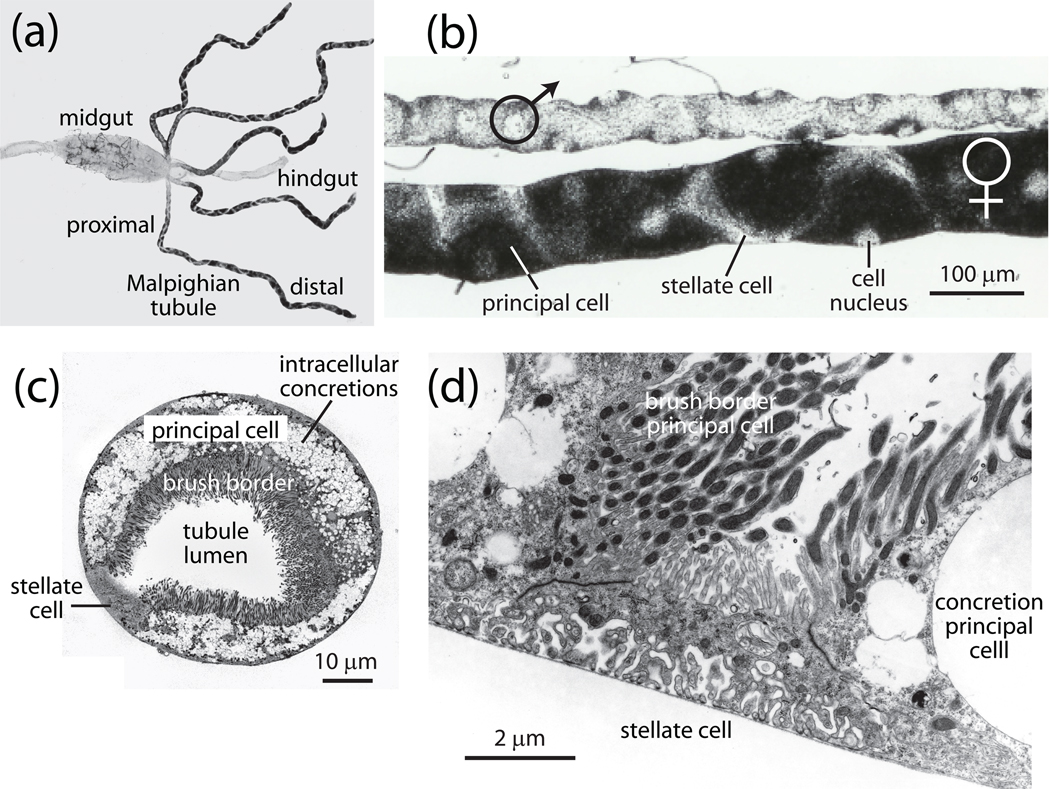
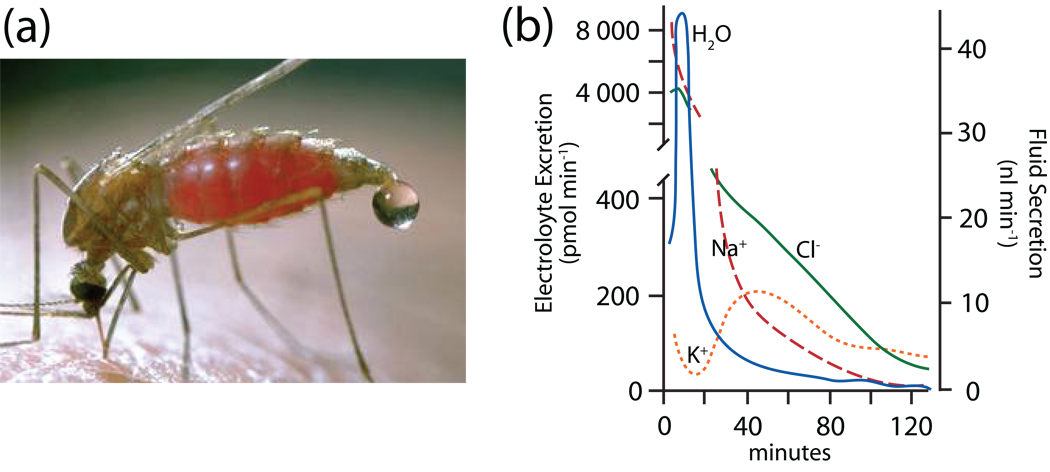
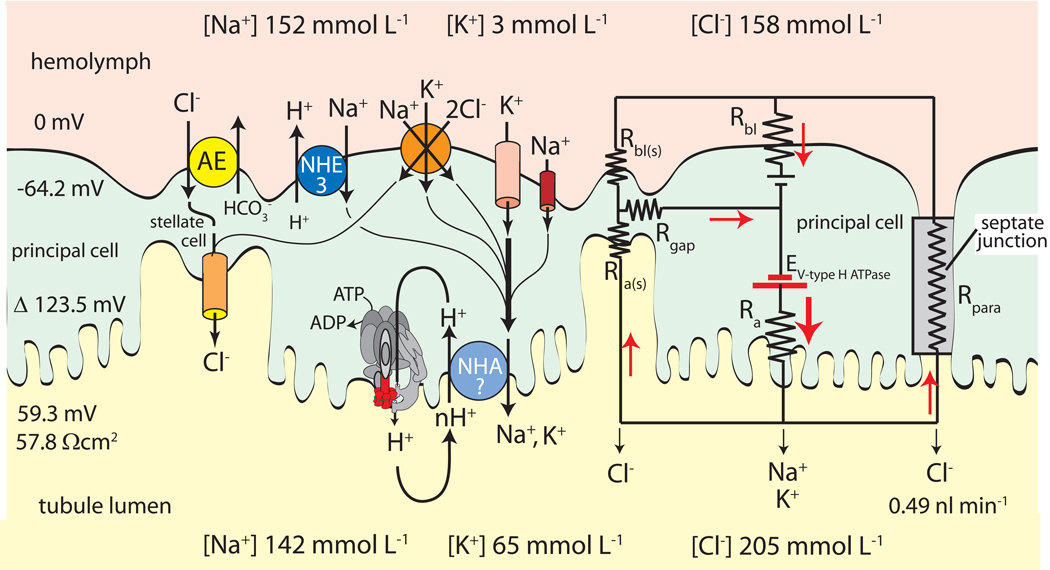
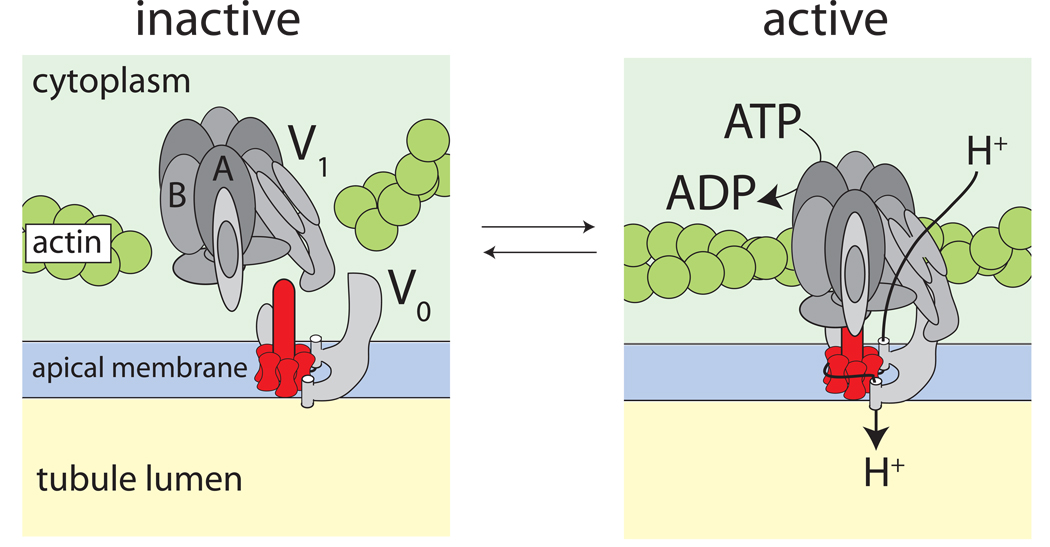
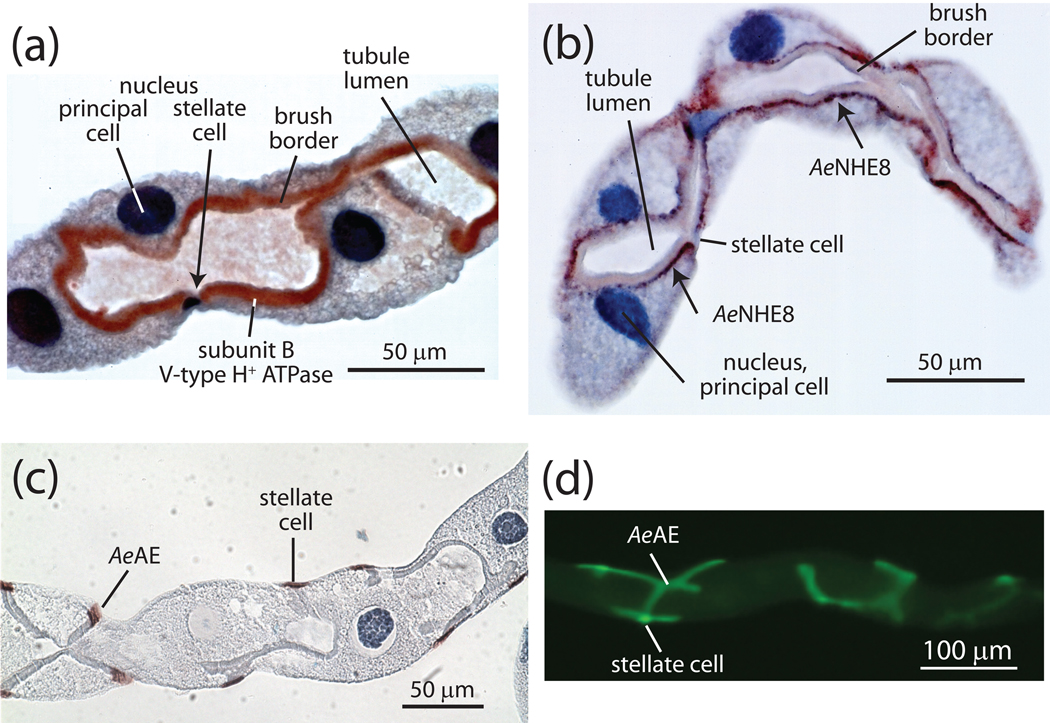
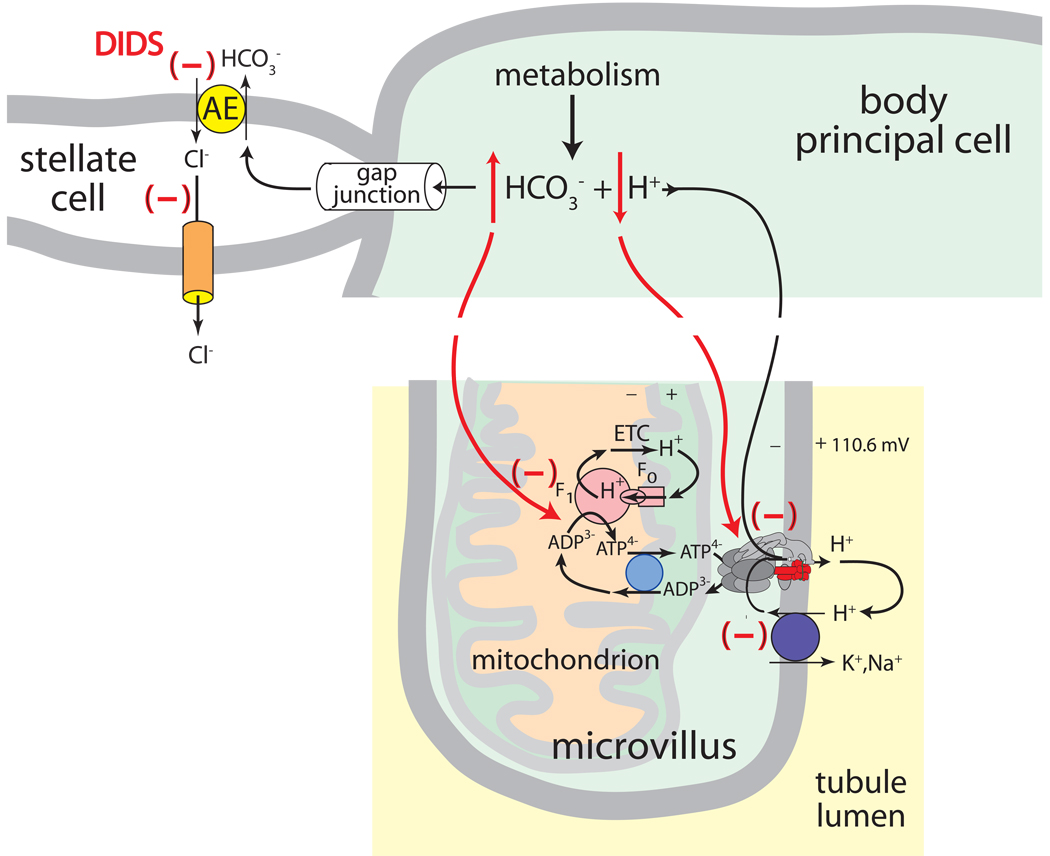
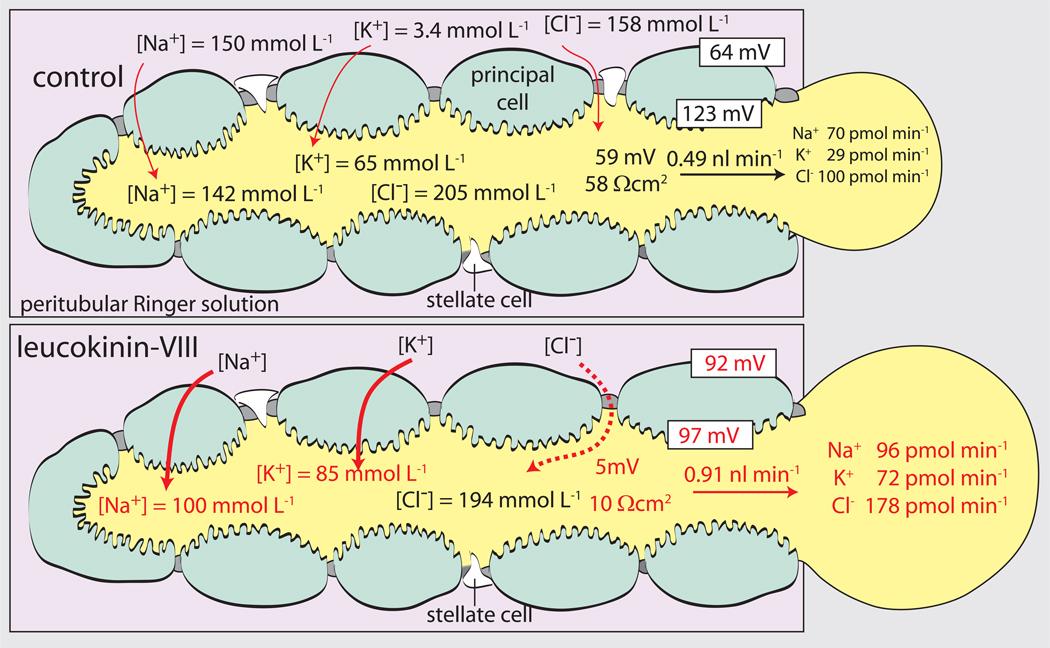
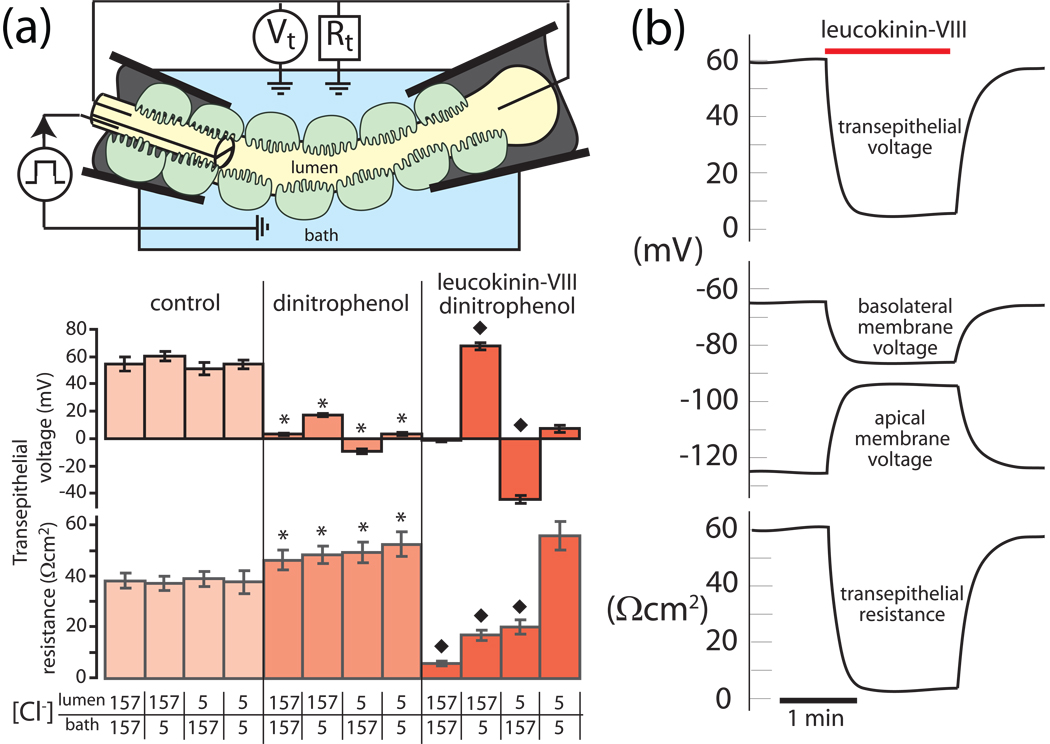
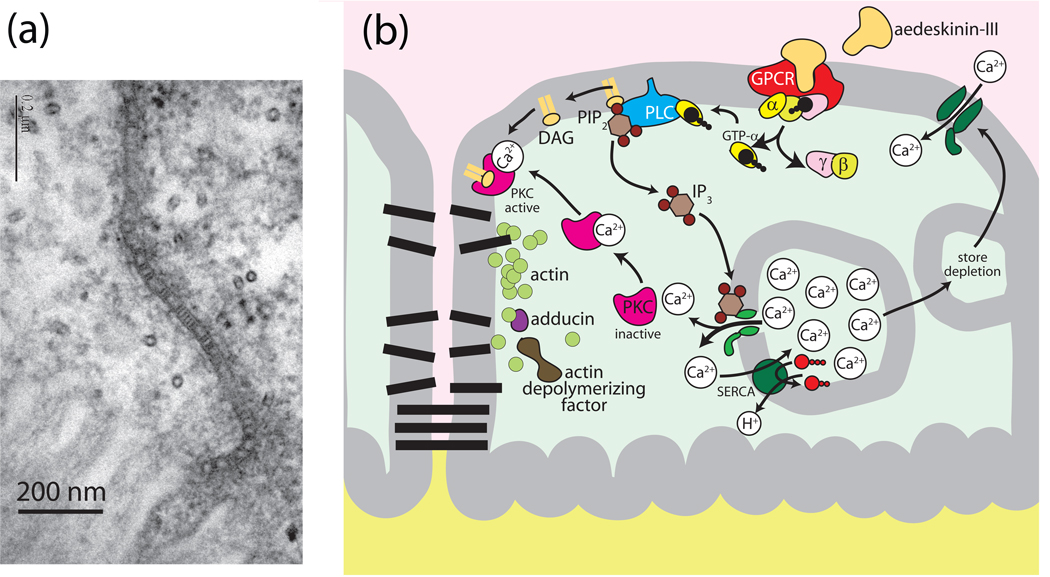
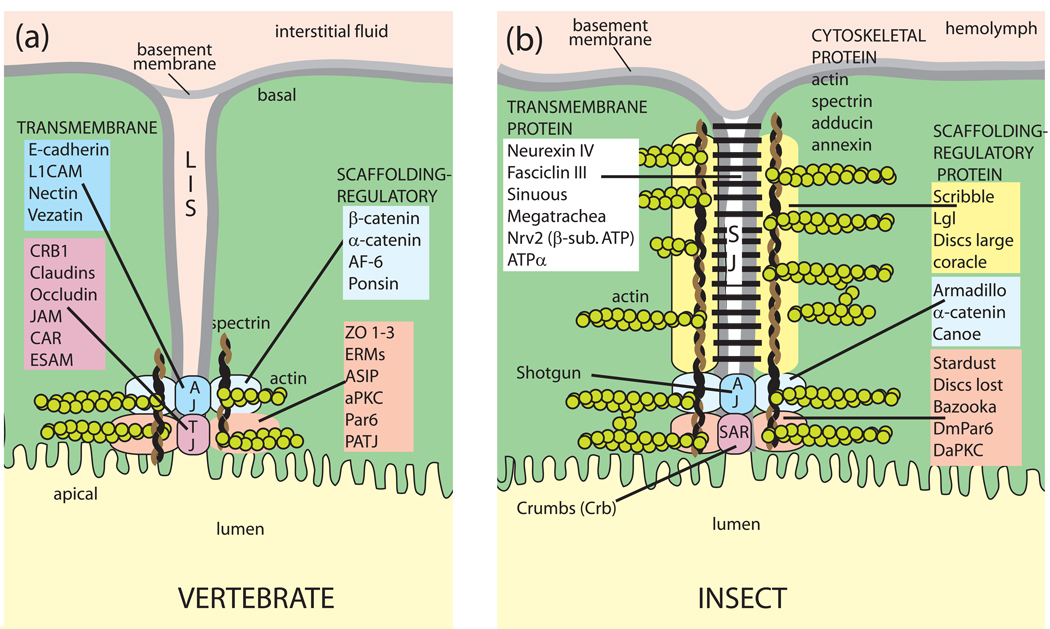
Isolated Malpighian tubules of the yellow fever mosquito secrete NaCl and KCl from the peritubular bath to the tubule lumen via active transport of Na(+) and K(+) by principal cells. Lumen-positive transepithelial voltages are the result. The counter-ion Cl(-) follows passively by electrodiffusion through the paracellular pathway. Water follows by osmosis, but specific routes for water across the epithelium are unknown. Remarkably, the transepithelial secretion of NaCl, KCl and water is driven by a H(+) V-ATPase located in the apical brush border membrane of principal cells and not the canonical Na(+), K(+) -ATPase. A hypothetical cation/H(+) exchanger moves Na(+) and K(+) from the cytoplasm to the tubule lumen. Also remarkable is the dynamic regulation of the paracellular permeability with switch-like speed which mediates in part the post-blood-meal diuresis in mosquitoes. For example, the blood meal the female mosquito takes to nourish her eggs triggers the release of kinin diuretic peptides that (i) increases the Cl(-) conductance of the paracellular pathway and (ii) assembles V(1) and V(0) complexes to activate the H(+) V-ATPase and cation/H(+) exchange close by. Thus, transcellular and paracellular pathways are both stimulated to quickly rid the mosquito of the unwanted salts and water of the blood meal. Stellate cells of the tubule appear to serve a metabolic support role, exporting the HCO(3)(-) generated during stimulated transport activity. Septate junctions define the properties of the paracellular pathway in Malpighian tubules, but the proteins responsible for the permselectivity and barrier functions of the septate junction are unknown.
© 2010 The Authors. Acta Physiologica © 2010 Scandinavian Physiological Society.
Figures
References
-
- Ahearn GA, Mandal PK, Mandal A. Biology of the 2Na/1H antiporter in invertebrates. J. exp. Biol. 2001;289:232–244. - PubMed
-
- Anderson JM, Van Itallie CM. Tight junctions. Curr Biol. 2008;18:R941–R943. - PubMed
-
- Bennett AF. Experimental evolution and the Krogh Principle: generating biological novelty for functional and genetic analyses. Physiol. Biochem. Zool. 2003;76:1–11. - PubMed
-
- Berg KA, Clarke WP. Development of functionally selective agonists as novel therapeutic agents. Drug Discov Today Ther Strateg. 2006;3:421–428.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
