

Patronin regulates the microtubule network by protecting microtubule minus ends
- PMID: 20946984
- PMCID: PMC3008421
- DOI: 10.1016/j.cell.2010.09.022
Patronin regulates the microtubule network by protecting microtubule minus ends
Abstract
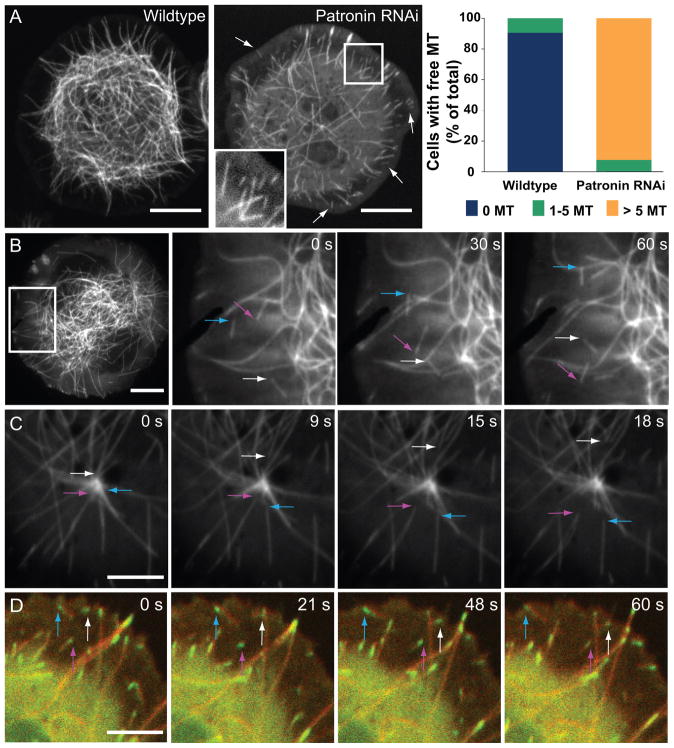
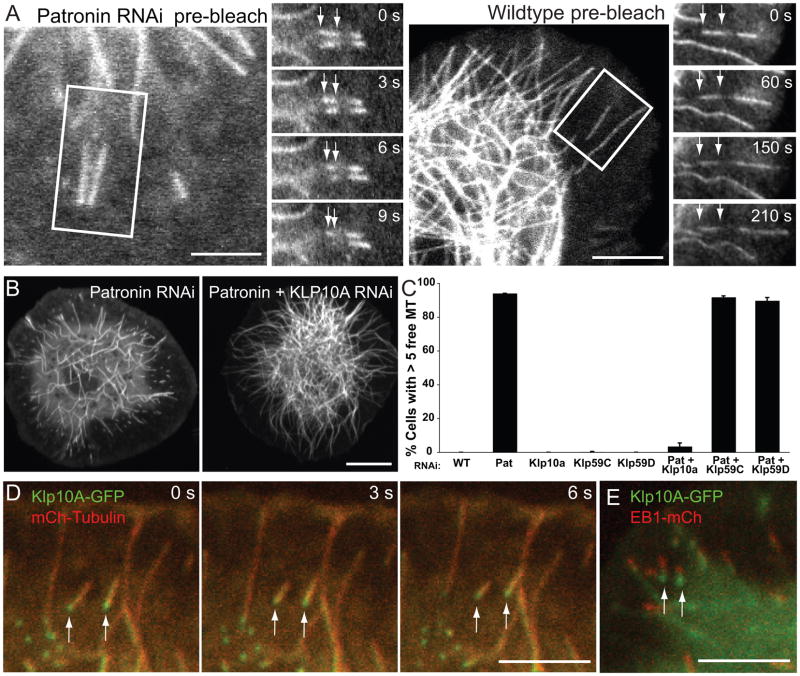
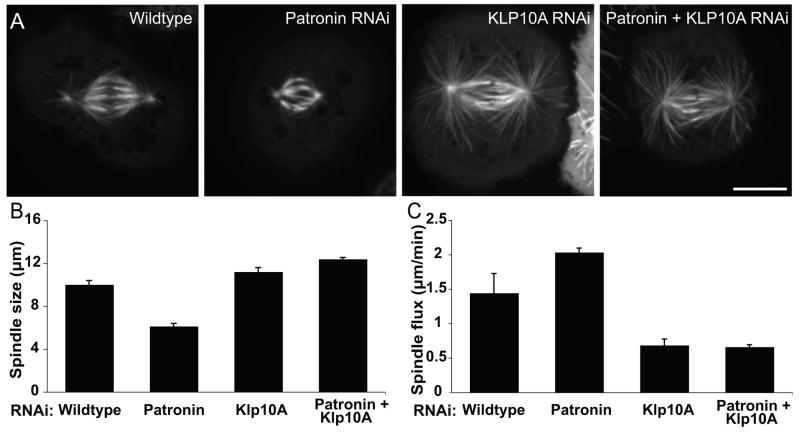
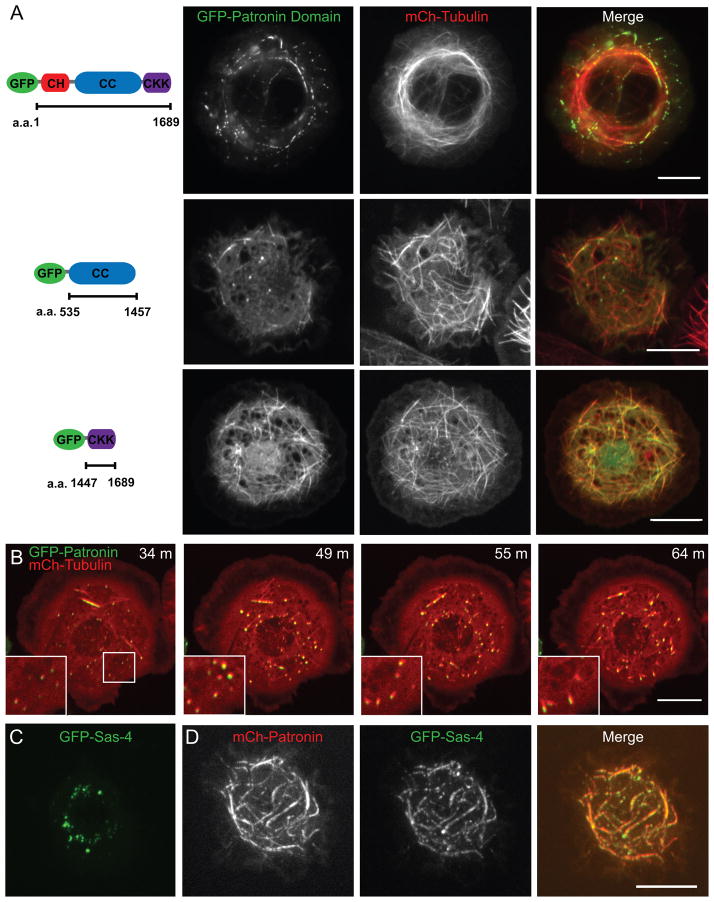
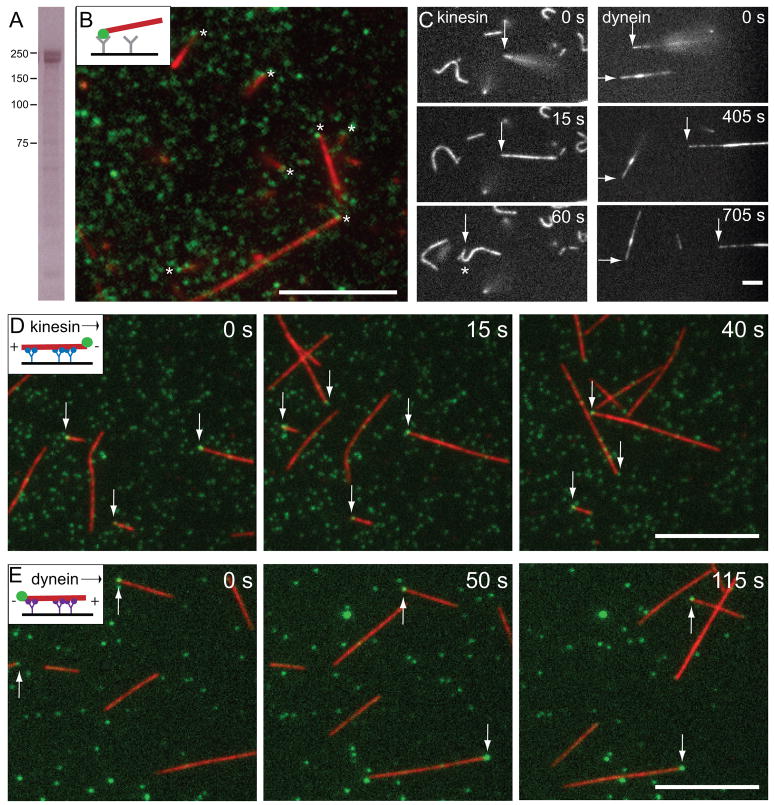
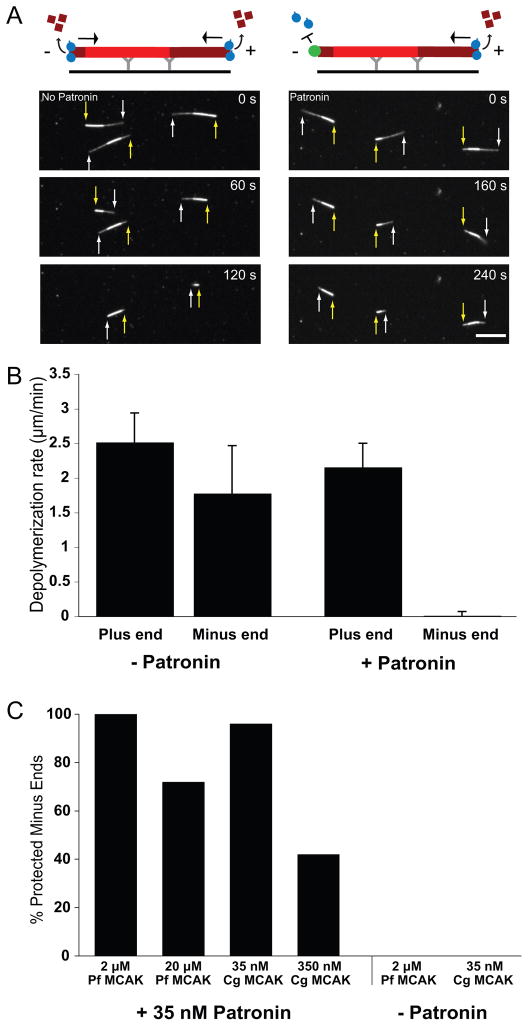
Tubulin assembles into microtubule polymers that have distinct plus and minus ends. Most microtubule plus ends in living cells are dynamic; the transitions between growth and shrinkage are regulated by assembly-promoting and destabilizing proteins. In contrast, minus ends are generally not dynamic, suggesting their stabilization by some unknown protein. Here, we have identified Patronin (also known as ssp4) as a protein that stabilizes microtubule minus ends in Drosophila S2 cells. In the absence of Patronin, minus ends lose subunits through the actions of the Kinesin-13 microtubule depolymerase, leading to a sparse interphase microtubule array and short, disorganized mitotic spindles. In vitro, the selective binding of purified Patronin to microtubule minus ends is sufficient to protect them against Kinesin-13-induced depolymerization. We propose that Patronin caps and stabilizes microtubule minus ends, an activity that serves a critical role in the organization of the microtubule cytoskeleton.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Cytoskeleton: Keeping minus ends stable.Nat Rev Mol Cell Biol. 2010 Dec;11(12):816. doi: 10.1038/nrm3014. Epub 2010 Nov 11. Nat Rev Mol Cell Biol. 2010. PMID: 21068769 No abstract available.
References
-
- Akhmanova A, Steinmetz M. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat Rev Mol Cell Biol. 2008;9:309–22. - PubMed
-
- Baines AJ, Bignone PA, King MD, Maggs AM, Bennett PM, Pinder JC, Phillips GW. The CKK domain (DUF1781) domain binds microtubules and defines the CAMSAP/ssp4 family of animal proteins. Mol Biol Evol. 2009;26:2005–14. - PubMed
-
- Bartolini F, Gundersen GG. Generation of noncentrosomal microtubule arrays. J Cell Sci. 2006;119:4155–63. - PubMed
-
- Berglund L, Björling E, Oksvold P, Fagerberg L, Asplund A, Al-Khalili Szigyarto C, Persson A, Ottosson J, Wernérus H, Nilsson P, et al. A genecentric Human Protein Atlas for expression profiles based on antibodies. Mol Cell Proteomics. 2008;7:2019–27. - PubMed
-
- Bornens M. Centrosome composition and microtubule anchoring mechanisms. Curr Opin Cell Biol. 2002;14:25–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
