

Identification of microRNAs regulating reprogramming factor LIN28 in embryonic stem cells and cancer cells
- PMID: 20947512
- PMCID: PMC3009922
- DOI: 10.1074/jbc.M110.169607
Identification of microRNAs regulating reprogramming factor LIN28 in embryonic stem cells and cancer cells
Abstract
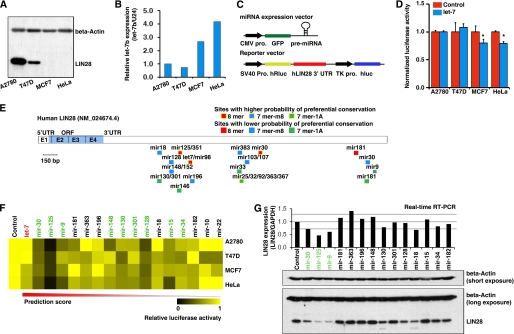
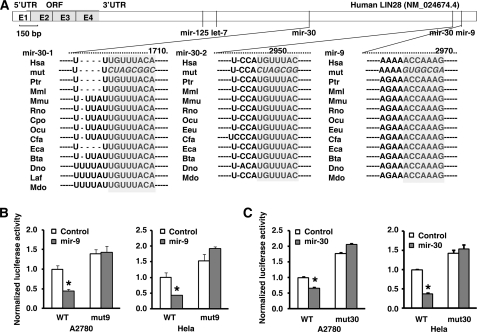
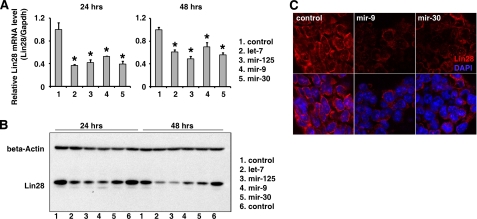
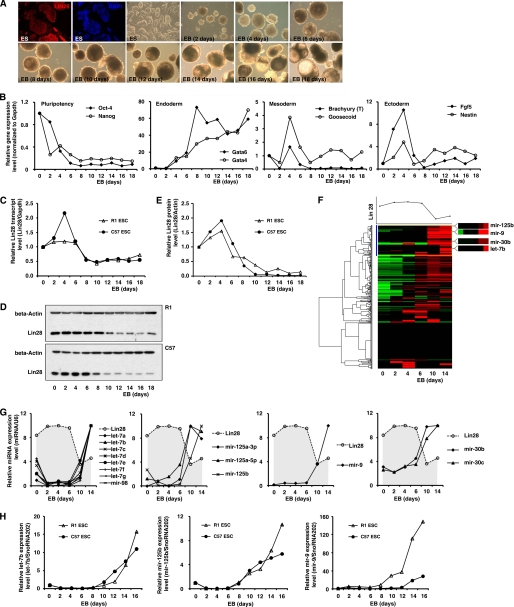
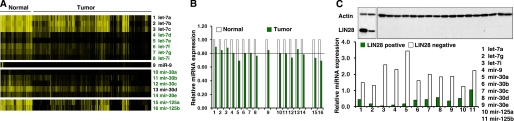
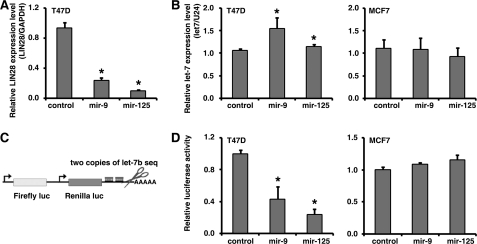
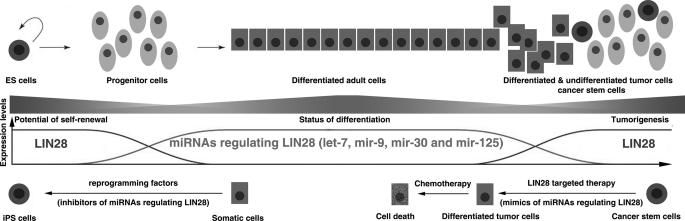
LIN28 (a homologue of the Caenorhabditis elegans lin-28 gene) is an evolutionarily conserved RNA-binding protein and a master regulator controlling the pluripotency of embryonic stem cells. Together with OCT4, SOX2, and NANOG, LIN28 can reprogram somatic cells, producing induced pluripotent stem cells. Expression of LIN28 is highly restricted to embryonic stem cells and developing tissues. In human tumors, LIN28 is up-regulated and functions as an oncogene promoting malignant transformation and tumor progression. However, the mechanisms of transcriptional and post-transcriptional regulation of LIN28 are still largely unknown. To examine microRNAs (miRNAs) that repress LIN28 expression, a combined in silico prediction and miRNA library screening approach was used in the present study. Four miRNAs directly regulating LIN28 (let-7, mir-125, mir-9, and mir-30) were initially identified by this approach and further validated by quantitative RT-PCR, Western blot analysis, and a LIN28 3'-UTR reporter assay. We found that expression levels of these four miRNAs were clustered together and inversely correlated with LIN28 expression during embryonic stem cell differentiation. In addition, the expression of these miRNAs was remarkably lower in LIN28-positive tumor cells compared with LIN28-negative tumor cells. Importantly, we demonstrated that these miRNAs were able to regulate the expression and activity of let-7, mediated by LIN28. Taken together, our studies demonstrate that miRNAs let-7, mir-125, mir-9, and mir-30 directly repress LIN28 expression in embryonic stem and cancer cells. Global down-regulation of these miRNAs may be one of the mechanisms of LIN28 reactivation in human cancers.
Figures
Similar articles
-
Embryonic stem cell microRNAs: defining factors in induced pluripotent (iPS) and cancer (CSC) stem cells?Curr Stem Cell Res Ther. 2009 Sep;4(3):168-77. doi: 10.2174/157488809789057400. Curr Stem Cell Res Ther. 2009. PMID: 19492978 Review.
-
Selective blockade of microRNA processing by Lin28.Science. 2008 Apr 4;320(5872):97-100. doi: 10.1126/science.1154040. Epub 2008 Feb 21. Science. 2008. PMID: 18292307 Free PMC article.
-
MicroRNA-145 Regulates Neural Stem Cell Differentiation Through the Sox2-Lin28/let-7 Signaling Pathway.Stem Cells. 2016 May;34(5):1386-95. doi: 10.1002/stem.2309. Epub 2016 Feb 29. Stem Cells. 2016. PMID: 26849971
-
miR-125b promotes early germ layer specification through Lin28/let-7d and preferential differentiation of mesoderm in human embryonic stem cells.PLoS One. 2012;7(4):e36121. doi: 10.1371/journal.pone.0036121. Epub 2012 Apr 24. PLoS One. 2012. PMID: 22545159 Free PMC article.
-
A mirror of two faces: Lin28 as a master regulator of both miRNA and mRNA.Wiley Interdiscip Rev RNA. 2012 Jul-Aug;3(4):483-94. doi: 10.1002/wrna.1112. Epub 2012 Mar 29. Wiley Interdiscip Rev RNA. 2012. PMID: 22467269 Review.
Cited by
-
Ovarian cancer stem cells: a new target for cancer therapy.Biomed Res Int. 2013;2013:916819. doi: 10.1155/2013/916819. Epub 2013 Jan 30. Biomed Res Int. 2013. PMID: 23509802 Free PMC article. Review.
-
OstemiR: a novel panel of microRNA biomarkers in osteoblastic and osteocytic differentiation from mesencymal stem cells.PLoS One. 2013;8(3):e58796. doi: 10.1371/journal.pone.0058796. Epub 2013 Mar 22. PLoS One. 2013. PMID: 23533592 Free PMC article.
-
Simulated Microgravity Exerts an Age-Dependent Effect on the Differentiation of Cardiovascular Progenitors Isolated from the Human Heart.PLoS One. 2015 Jul 10;10(7):e0132378. doi: 10.1371/journal.pone.0132378. eCollection 2015. PLoS One. 2015. PMID: 26161778 Free PMC article.
-
Human Induced Pluripotent Stem Cell-Derived Vascular Cells: Recent Progress and Future Directions.J Cardiovasc Dev Dis. 2021 Nov 4;8(11):148. doi: 10.3390/jcdd8110148. J Cardiovasc Dev Dis. 2021. PMID: 34821701 Free PMC article. Review.
-
Role of LIN28A in mouse and human trophoblast cell differentiation.Biol Reprod. 2013 Oct 24;89(4):95. doi: 10.1095/biolreprod.113.109868. Print 2013 Oct. Biol Reprod. 2013. PMID: 24006280 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
