

Germline KRAS mutations cause aberrant biochemical and physical properties leading to developmental disorders
- PMID: 20949621
- PMCID: PMC3117284
- DOI: 10.1002/humu.21377
Germline KRAS mutations cause aberrant biochemical and physical properties leading to developmental disorders
Abstract
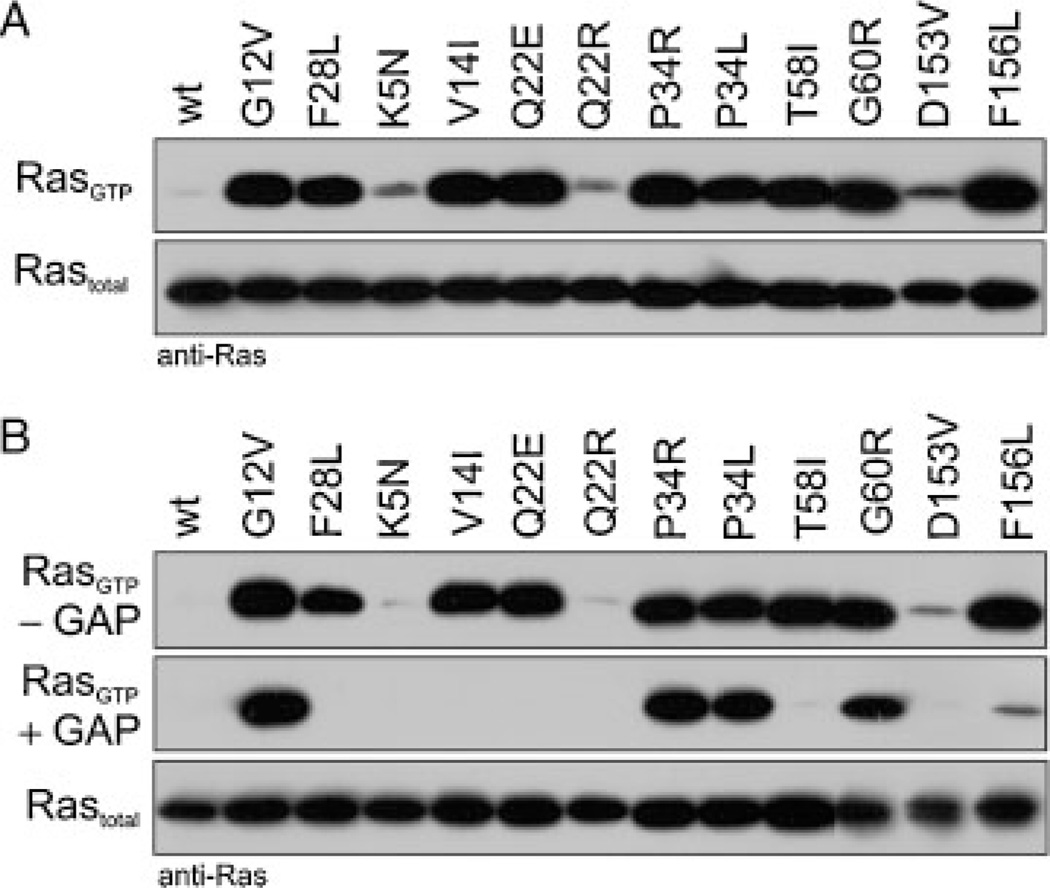
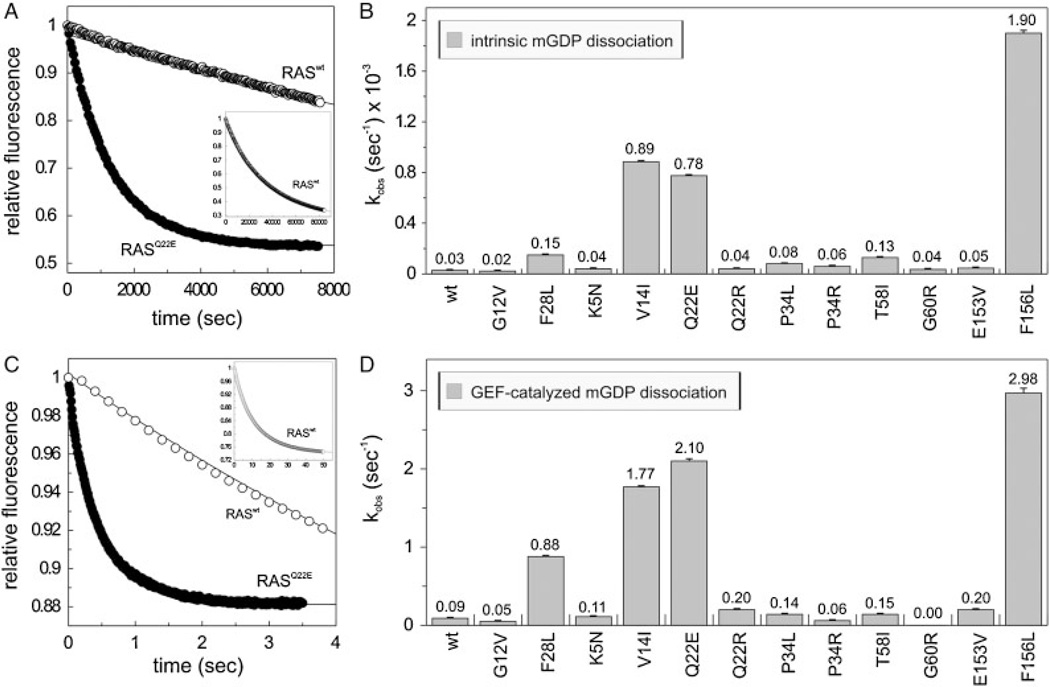
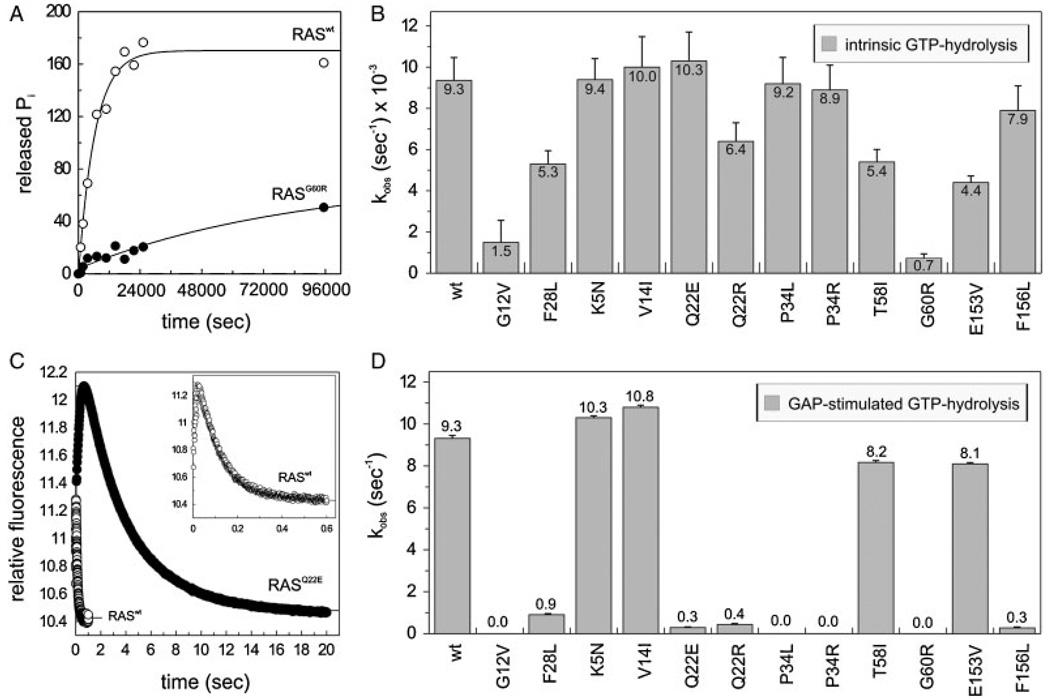
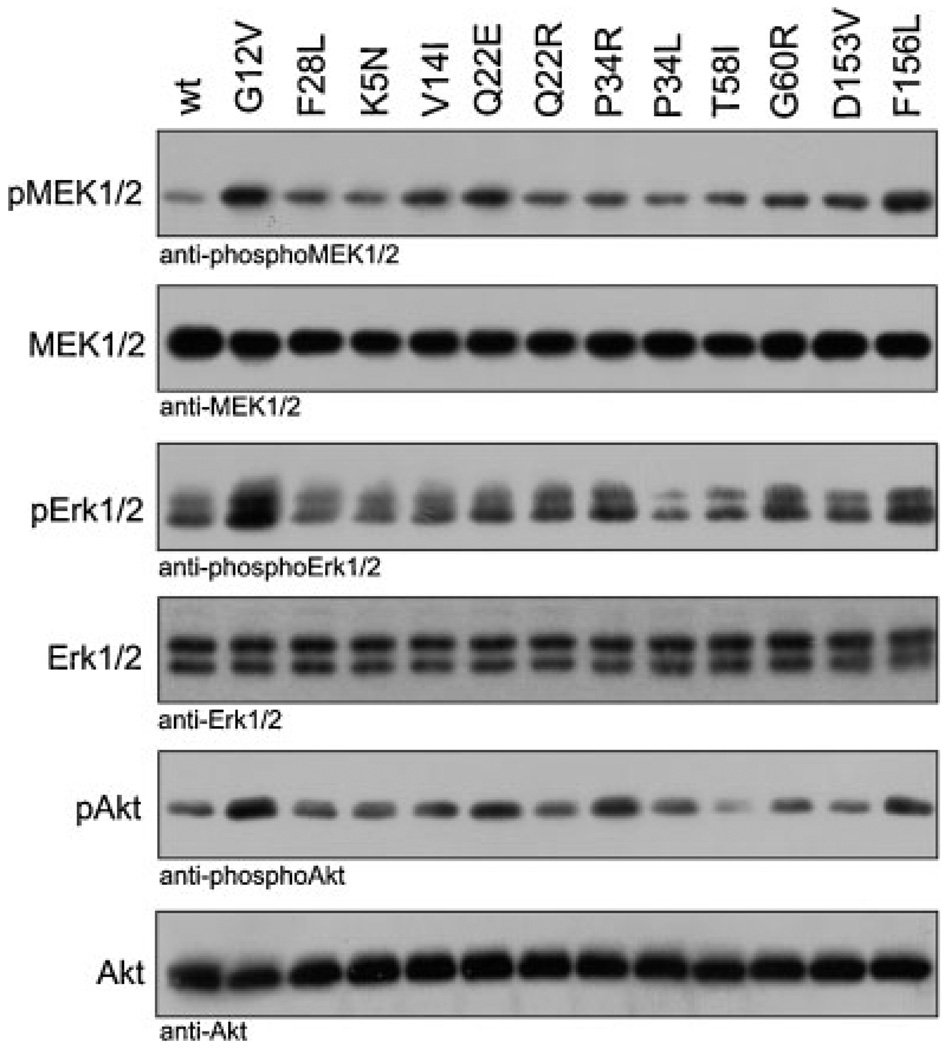
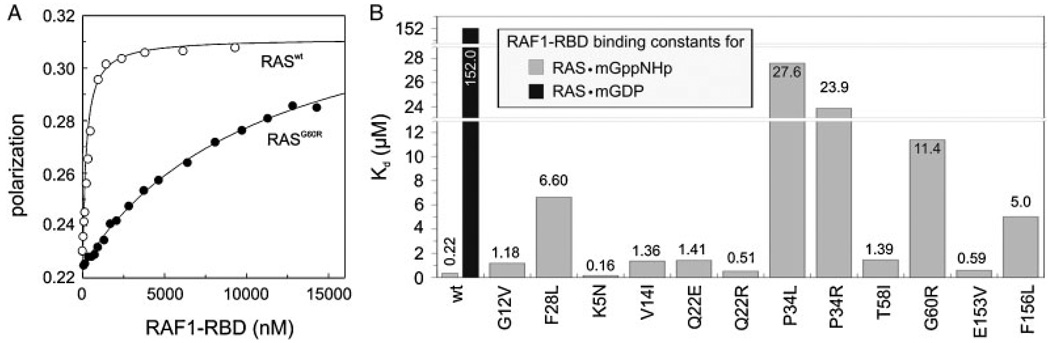
The KRAS gene is the most common locus for somatic gain-of-function mutations in human cancer. Germline KRAS mutations were shown recently to be associated with developmental disorders, including Noonan syndrome (NS), cardio-facio-cutaneous syndrome (CFCS), and Costello syndrome (CS). The molecular basis of this broad phenotypic variability has in part remained elusive so far. Here, we comprehensively analyzed the biochemical and structural features of ten germline KRAS mutations using physical and cellular biochemistry. According to their distinct biochemical and structural alterations, the mutants can be grouped into five distinct classes, four of which markedly differ from RAS oncoproteins. Investigated functional alterations comprise the enhancement of intrinsic and guanine nucleotide exchange factor (GEF) catalyzed nucleotide exchange, which is alternatively accompanied by an impaired GTPase-activating protein (GAP) stimulated GTP hydrolysis, an overall loss of functional properties, and a deficiency in effector interaction. In conclusion, our data underscore the important role of RAS in the pathogenesis of the group of related disorders including NS, CFCS, and CS, and provide clues to the high phenotypic variability of patients with germline KRAS mutations.
© 2010 Wiley-Liss, Inc.
Figures

References
-
- Ahmadian MR. Prospects for anti-ras drugs. Br J Haematol. 2002;116:511–518. - PubMed
-
- Ahmadian MR, Hoffmann U, Goody RS, Wittinghofer A. Individual rate constants for the interaction of Ras proteins with GTPase-activating proteins determined by fluorescence spectroscopy. Biochemistry. 1997;36:4535–4541. - PubMed
-
- Ahmadian MR, Wittinghofer A, Herrmann C. Fluorescence methods in the study of small GTP-binding proteins. Methods Mol Biol. 2002;189:45–63. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
