

Long-term genetic stability and population dynamics of laboratory strains of Schistosoma mansoni
- PMID: 20950096
- PMCID: PMC3881424
- DOI: 10.1645/GE-2463.1
Long-term genetic stability and population dynamics of laboratory strains of Schistosoma mansoni
Abstract
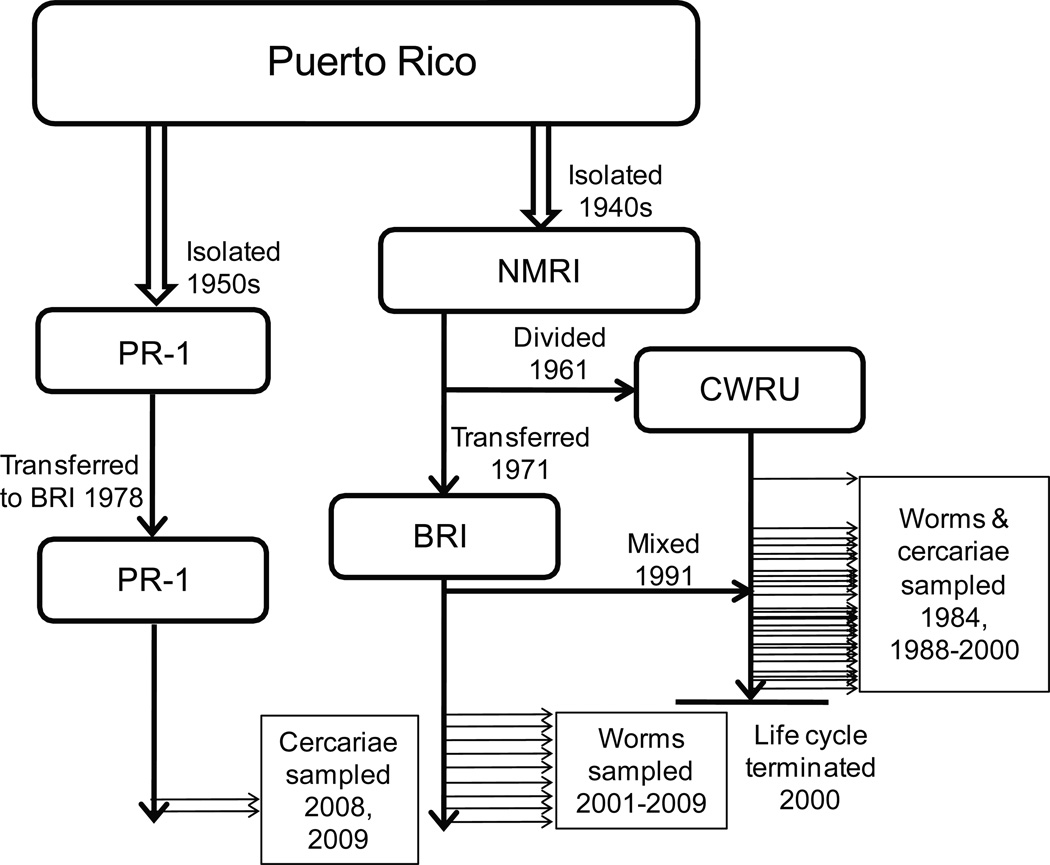
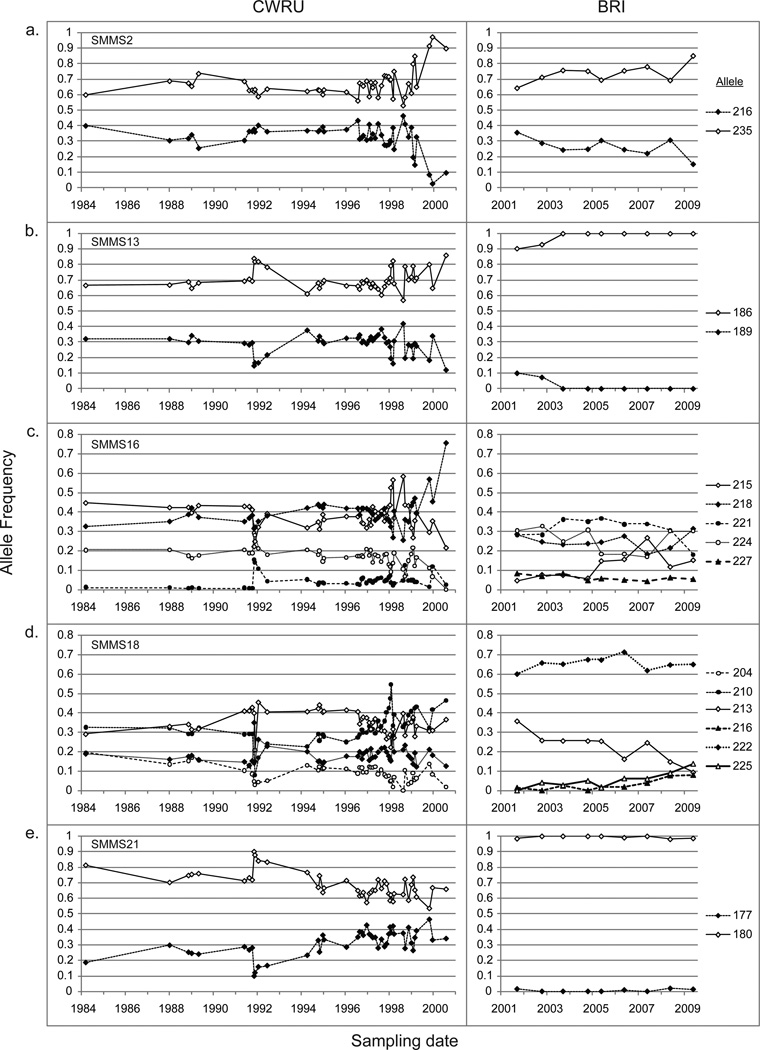
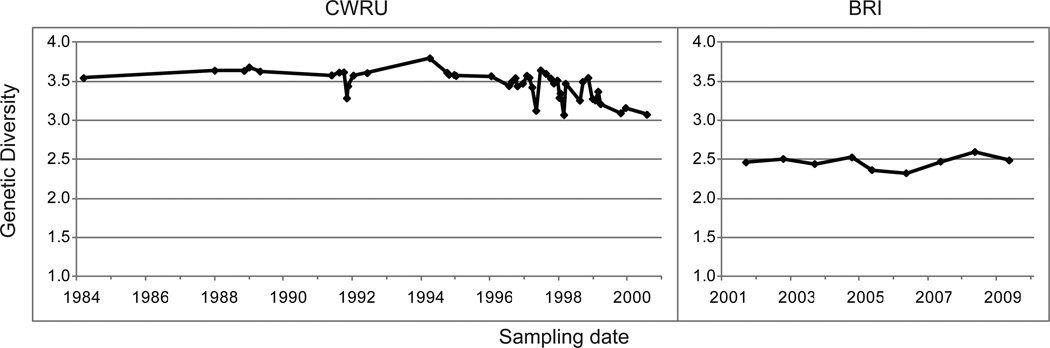
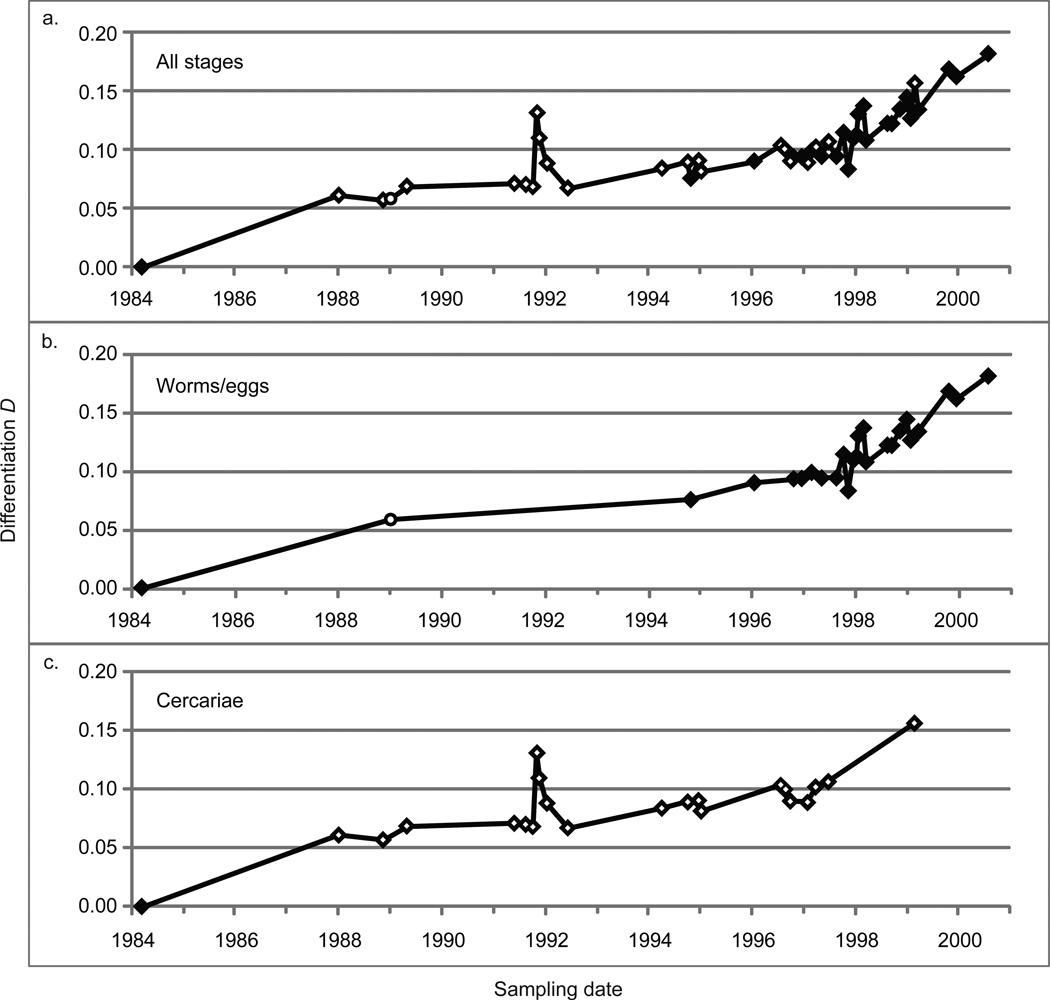
Measures of genetic differentiation between populations are useful tools for understanding the long-term dynamics of parasite communities. We followed the allele frequencies of microsatellite markers in samples taken over a period of 16 yr from the Case Western Reserve University-Naval Medical Research Institute (CWRU-NMRI) laboratory strain of Schistosoma mansoni. DNA was isolated from pooled samples of adults, eggs, or cercariae collected at 46 time points and genotyped for 14 tri- or tetranucleotide microsatellite markers. For comparison, 2 S. mansoni reference strains (Biomedical Research Institute-NMRI, which has a common origin with the CWRU line, and PR-1) were analyzed over shorter periods of time. We observed that the long-term allele frequencies are generally stable in large laboratory populations of this parasite, and a high degree of similarity was observed between the allele frequencies of consecutive samples from different developmental stages. The CWRU strain, however, showed 2 periods of marked deviation from stability as demonstrated using genetic differentiation measures. The first period corresponds to an admixture event with the BRI strain in which a new equilibrium was established as the "migrants" became blended into the existing CWRU population, consistent with 23% admixture from BRI. The second corresponds to a period of genetic drift when the CWRU population size was greatly reduced with an accompanying loss in genetic diversity. Having demonstrated the utility of pooled samples for the genetic analysis of population dynamics in laboratory strains of schistosomes, this approach will be useful for analyzing field samples to determine the impact of schistosomiasis control programs on parasite population structure. Accounting only for the intensity or prevalence of parasite populations may fail to register significant changes in population structure that could have implications for resistance, morbidity, and the design of control measures.
Figures

References
-
- Chao A, Shen TJ. SPADE (Species Prediction And Diversity Estimation) 2003 Program and User’s Guide published at http://chao.stat.nthu.edu.tw.
-
- Chao A, Jost L, Chiang SC, Jiang YH, Chazdon RL. A two-stage probabilistic approach to multiple-community similarity indices. Biometrics. 2008;64:1178–1186. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
