

Mitochondrial involvement in cell death of non-mammalian eukaryotes
- PMID: 20950655
- PMCID: PMC3033473
- DOI: 10.1016/j.bbamcr.2010.10.008
Mitochondrial involvement in cell death of non-mammalian eukaryotes
Abstract
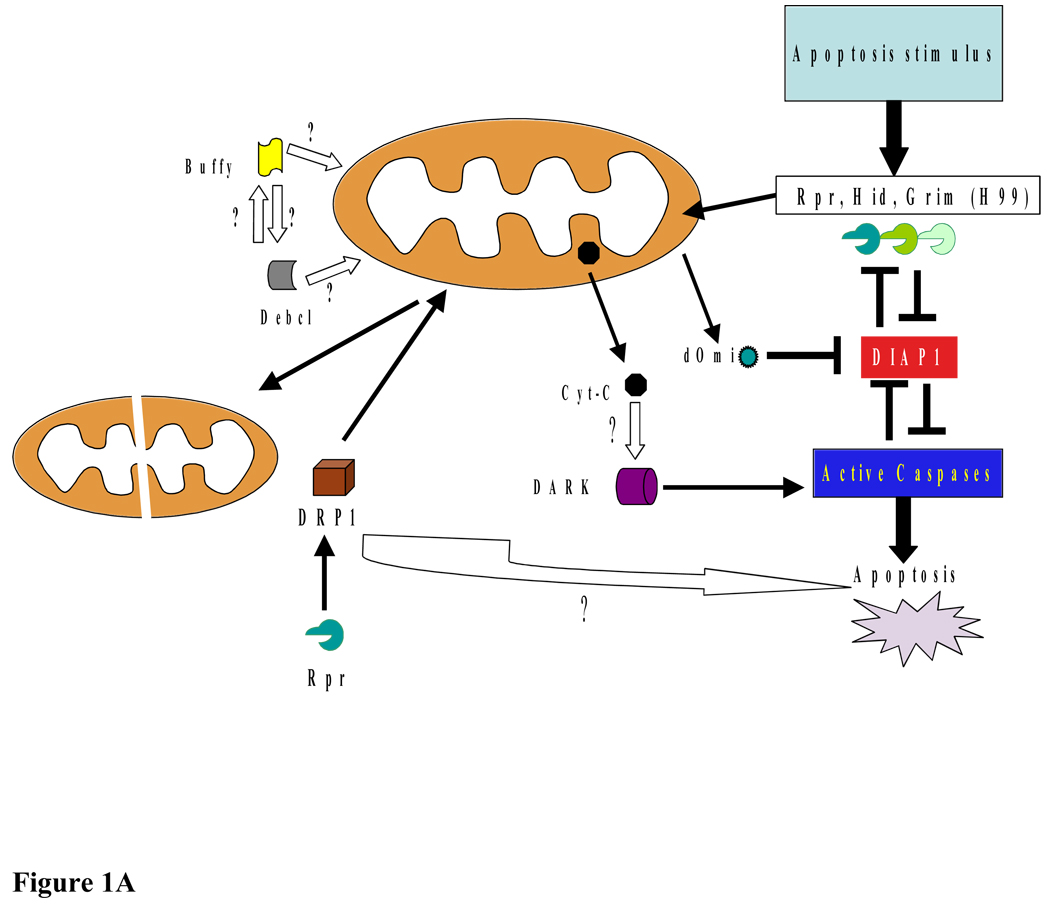
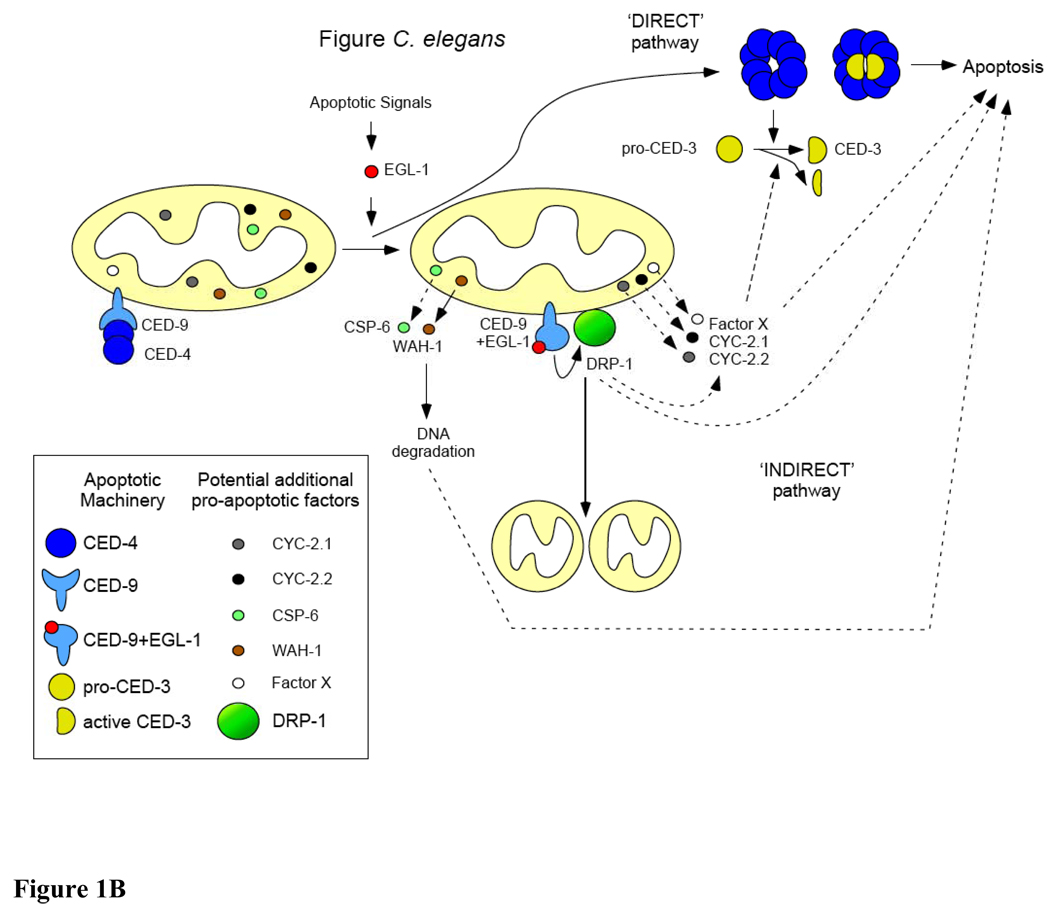
Although mitochondria are essential organelles for long-term survival of eukaryotic cells, recent discoveries in biochemistry and genetics have advanced our understanding of the requirements for mitochondria in cell death. Much of what we understand about cell death is based on the identification of conserved cell death genes in Drosophila melanogaster and Caenorhabditis elegans. However, the role of mitochondria in cell death in these models has been much less clear. Considering the active role that mitochondria play in apoptosis in mammalian cells, the mitochondrial contribution to cell death in non-mammalian systems has been an area of active investigation. In this article, we review the current research on this topic in three non-mammalian models, C. elegans, Drosophila, and Saccharomyces cerevisiae. In addition, we discuss how non-mammalian models have provided important insight into the mechanisms of human disease as they relate to the mitochondrial pathway of cell death. The unique perspective derived from each of these model systems provides a more complete understanding of mitochondria in programmed cell death. This article is part of a Special Issue entitled Mitochondria: the deadly organelle.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
References
-
- Yuan J, Shaham S, Ledoux S, Ellis HM, Horvitz HR. The C. elegans cell death gene ced-3 encodes a protein similar to mammalian interleukin-1 beta-converting enzyme. Cell. 1993;75:641–652. - PubMed
-
- Miura M, Zhu H, Rotello R, Hartwieg EA, Yuan J. Induction of apoptosis in fibroblasts by IL-1 beta-converting enzyme, a mammalian homolog of the C. elegans cell death gene ced-3. Cell. 1993;75:653–660. - PubMed
-
- Hengartner MO, Horvitz HR. C. elegans cell survival gene ced-9 encodes a functional homolog of the mammalian proto-oncogene bcl-2. 1994;76:665–676. - PubMed
-
- Hay BA, Wassarman DA, Rubin GM. Drosophila homologs of baculovirus inhibitor of apoptosis proteins function to block cell death. Cell. 1995;83:1253–1262. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
