

Binocular vision
- PMID: 20951722
- PMCID: PMC3050089
- DOI: 10.1016/j.visres.2010.10.009
Binocular vision
Abstract
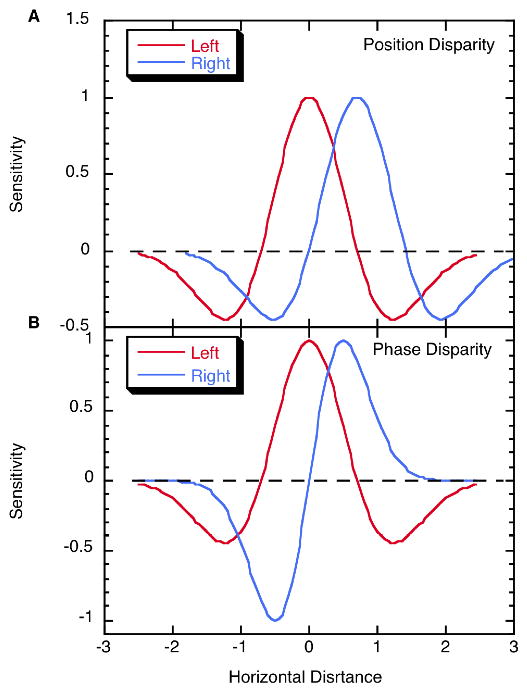
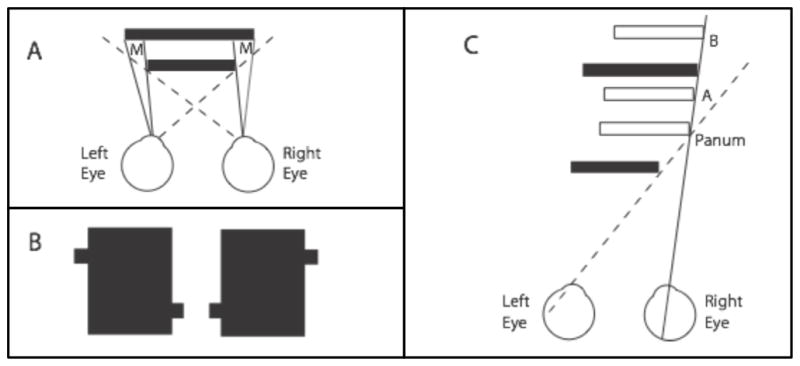
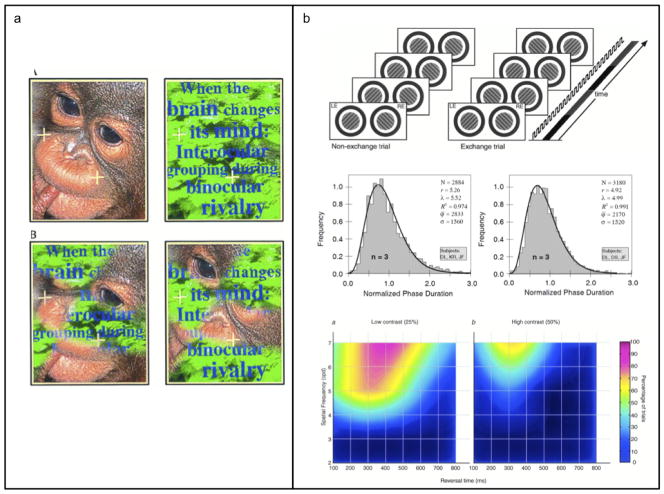
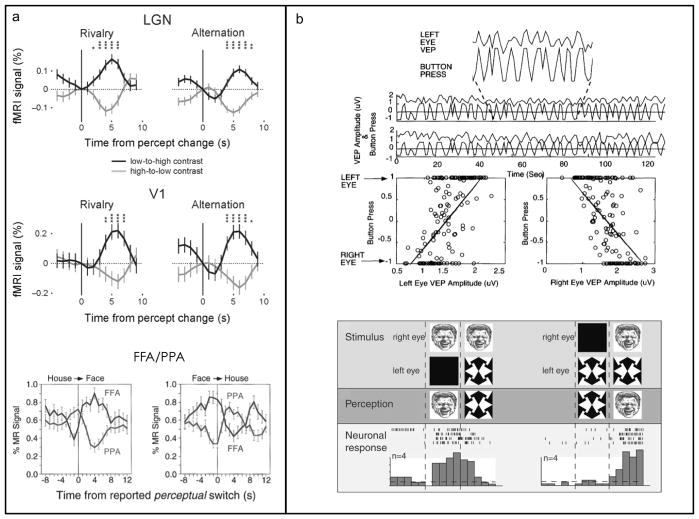
This essay reviews major developments - empirical and theoretical - in the field of binocular vision during the last 25years. We limit our survey primarily to work on human stereopsis, binocular rivalry and binocular contrast summation, with discussion where relevant of single-unit neurophysiology and human brain imaging. We identify several key controversies that have stimulated important work on these problems. In the case of stereopsis those controversies include position vs. phase encoding of disparity, dependence of disparity limits on spatial scale, role of occlusion in binocular depth and surface perception, and motion in 3D. In the case of binocular rivalry, controversies include eye vs. stimulus rivalry, role of "top-down" influences on rivalry dynamics, and the interaction of binocular rivalry and stereopsis. Concerning binocular contrast summation, the essay focuses on two representative models that highlight the evolving complexity in this field of study.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Adelson EH, Bergen JR. Spatiotemporal energy models for the perception of motion. Journal of the Optical Society of America, A. 1985;2:284–299. - PubMed
-
- Alais D, Blake R. Binocular rivalry and perceptual ambiguity. Boston: MIT Press; 2005.
-
- Alais D, Lorenceau J, Arrighi R, Cass JR. Contour interactions between pairs of Gabors engaged in binocular rivalry reveal a map of the association field. Vision Research. 2006;46:1473–1487. - PubMed
-
- Alais D, Melcher D. Strength and coherence of binocular rivalry depends on shared stimulus complexity. Vision Research. 2007;47:269–279. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
