

A phospho/methyl switch at histone H3 regulates TFIID association with mitotic chromosomes
- PMID: 20953165
- PMCID: PMC3020634
- DOI: 10.1038/emboj.2010.261
A phospho/methyl switch at histone H3 regulates TFIID association with mitotic chromosomes
Abstract
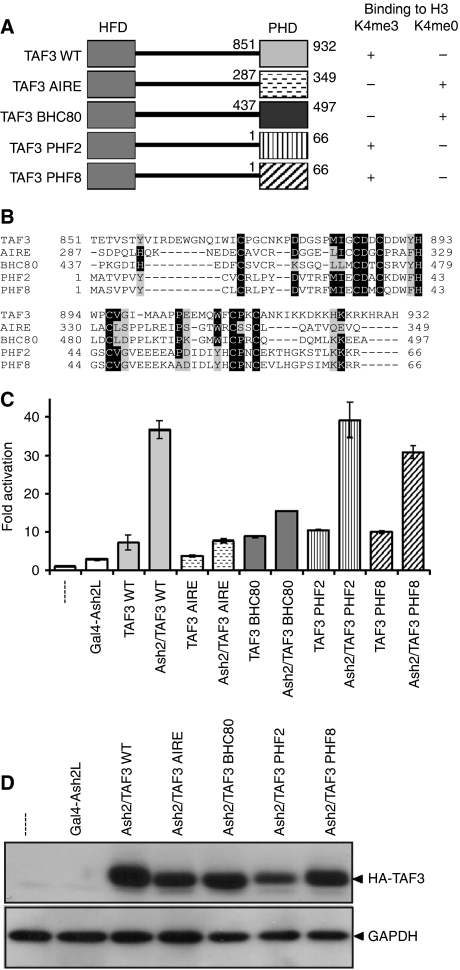
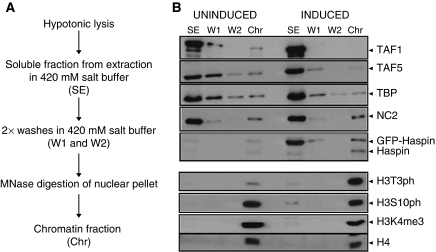
Histone methylation patterns are correlated with eukaryotic gene transcription. High-affinity binding of the plant homeodomain (PHD) of TFIID subunit TAF3 to trimethylated lysine-4 of histone H3 (H3K4me3) is involved in promoter recruitment of this basal transcription factor. Here, we show that for transcription activation the PHD of TAF3 can be replaced by PHDs of other high-affinity H3K4me3 binders. Interestingly, H3K4me3 binding of TFIID and the TAF3-PHD is decreased by phosphorylation of the adjacent threonine residue (H3T3), which coincides with mitotic inhibition of transcription. Ectopic expression of the H3T3 kinase haspin repressed TAF3-mediated transcription of endogenous and of reporter genes and decreased TFIID association with chromatin. Conversely, immunofluorescence and live-cell microscopy studies showed an increased association of TFIID with mitotic chromosomes upon haspin knockdown. Based on our observations, we propose that a histone H3 phospho-methyl switch regulates TFIID-mediated transcription during mitotic progression of the cell cycle.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





References
-
- Bernstein BE, Kamal M, Lindblad-Toh K, Bekiranov S, Bailey DK, Huebert DJ, McMahon S, Karlsson EK, Kulbokas EJ III, Gingeras TR, Schreiber SL, Lander ES (2005) Genomic maps and comparative analysis of histone modifications in human and mouse. Cell 120: 169–181 - PubMed
-
- Christova R, Oelgeschlager T (2002) Association of human TFIID-promoter complexes with silenced mitotic chromatin in vivo. Nat Cell Biol 4: 79–82 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
