

The program for processing newly synthesized histones H3.1 and H4
- PMID: 20953179
- PMCID: PMC2988979
- DOI: 10.1038/nsmb.1911
The program for processing newly synthesized histones H3.1 and H4
Abstract
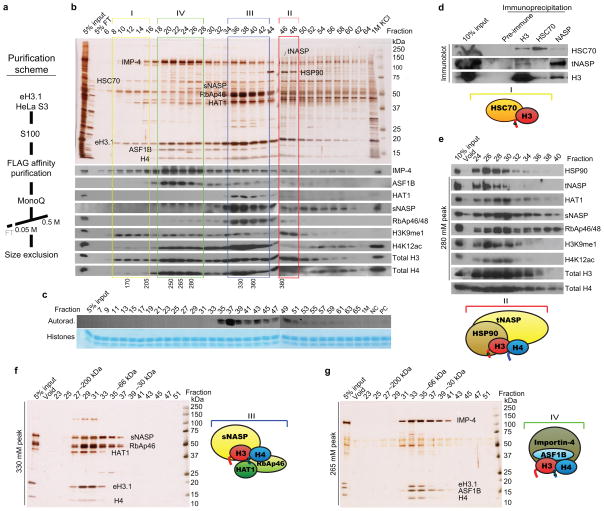
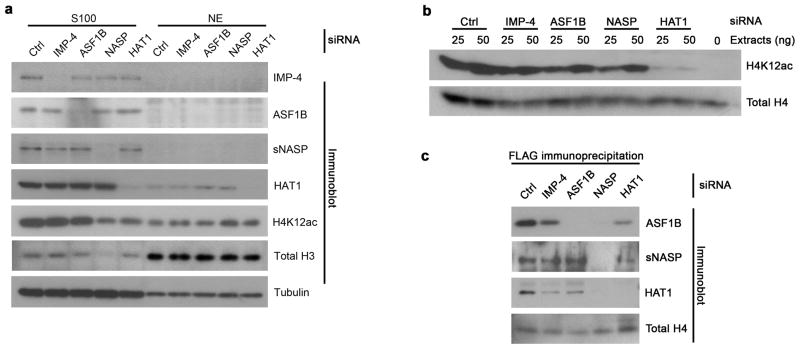
The mechanism by which newly synthesized histones are imported into the nucleus and deposited onto replicating chromatin alongside segregating nucleosomal counterparts is poorly understood, yet this program is expected to bear on the putative epigenetic nature of histone post-translational modifications. To define the events by which naive pre-deposition histones are imported into the nucleus, we biochemically purified and characterized the full gamut of histone H3.1-containing complexes from human cytoplasmic fractions and identified their associated histone post-translational modifications. Through reconstitution assays, biophysical analyses and live cell manipulations, we describe in detail this series of events, namely the assembly of H3-H4 dimers, the acetylation of histones by the HAT1 holoenzyme and the transfer of histones between chaperones that culminates with their karyopherin-mediated nuclear import. We further demonstrate the high degree of conservation for this pathway between higher and lower eukaryotes.
Figures





References
-
- Eickbush TH, Moudrianakis EN. The histone core complex: an octamer assembled by two sets of protein-protein interactions. Biochemistry. 1978;17:4955–64. - PubMed
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–60. - PubMed
-
- Campos EI, Reinberg D. Histones: Annotating Chromatin. Annual Review of Genetics. 2009;43:559–599. - PubMed
-
- Koessler H, Doenecke D, Albig W. Aberrant expression pattern of replication-dependent histone h3 subtype genes in human tumor cell lines. DNA Cell Biol. 2003;22:233–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
