

Neurophysiological origin of human brain asymmetry for speech and language
- PMID: 20956297
- PMCID: PMC2972980
- DOI: 10.1073/pnas.1007189107
Neurophysiological origin of human brain asymmetry for speech and language
Abstract
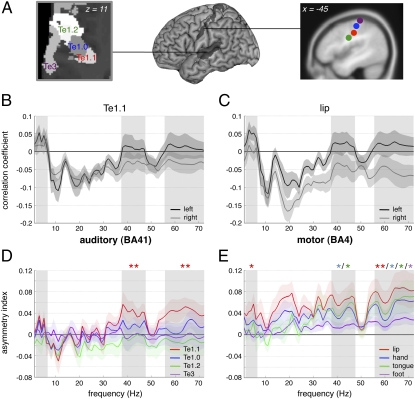
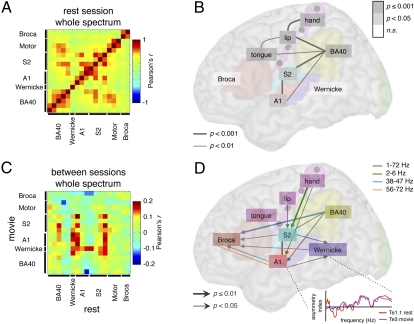
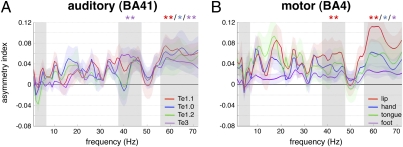
The physiological basis of human cerebral asymmetry for language remains mysterious. We have used simultaneous physiological and anatomical measurements to investigate the issue. Concentrating on neural oscillatory activity in speech-specific frequency bands and exploring interactions between gestural (motor) and auditory-evoked activity, we find, in the absence of language-related processing, that left auditory, somatosensory, articulatory motor, and inferior parietal cortices show specific, lateralized, speech-related physiological properties. With the addition of ecologically valid audiovisual stimulation, activity in auditory cortex synchronizes with left-dominant input from the motor cortex at frequencies corresponding to syllabic, but not phonemic, speech rhythms. Our results support theories of language lateralization that posit a major role for intrinsic, hardwired perceptuomotor processing in syllabic parsing and are compatible both with the evolutionary view that speech arose from a combination of syllable-sized vocalizations and meaningful hand gestures and with developmental observations suggesting phonemic analysis is a developmentally acquired process.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bailey WJ, Yang S. Hearing asymmetry and auditory acuity in the Australian bushcricket Requena verticalis (Listroscelidinae; Tettigoniidae; Orthoptera) J Exp Biol. 2002;205:2935–2942. - PubMed
-
- Meguerditchian A, Vauclair J. Contrast of hand preferences between communicative gestures and non-communicative actions in baboons: Implications for the origins of hemispheric specialization for language. Brain Lang. 2009;108:167–174. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
