

c-Src differentially regulates the functions of microtentacles and invadopodia
- PMID: 20956943
- PMCID: PMC4073667
- DOI: 10.1038/onc.2010.360
c-Src differentially regulates the functions of microtentacles and invadopodia
Abstract
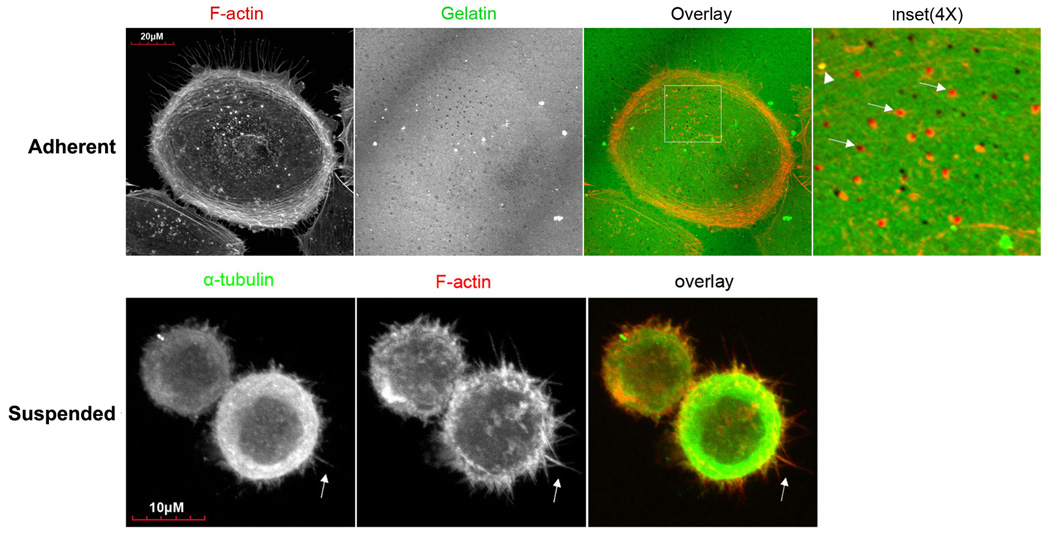
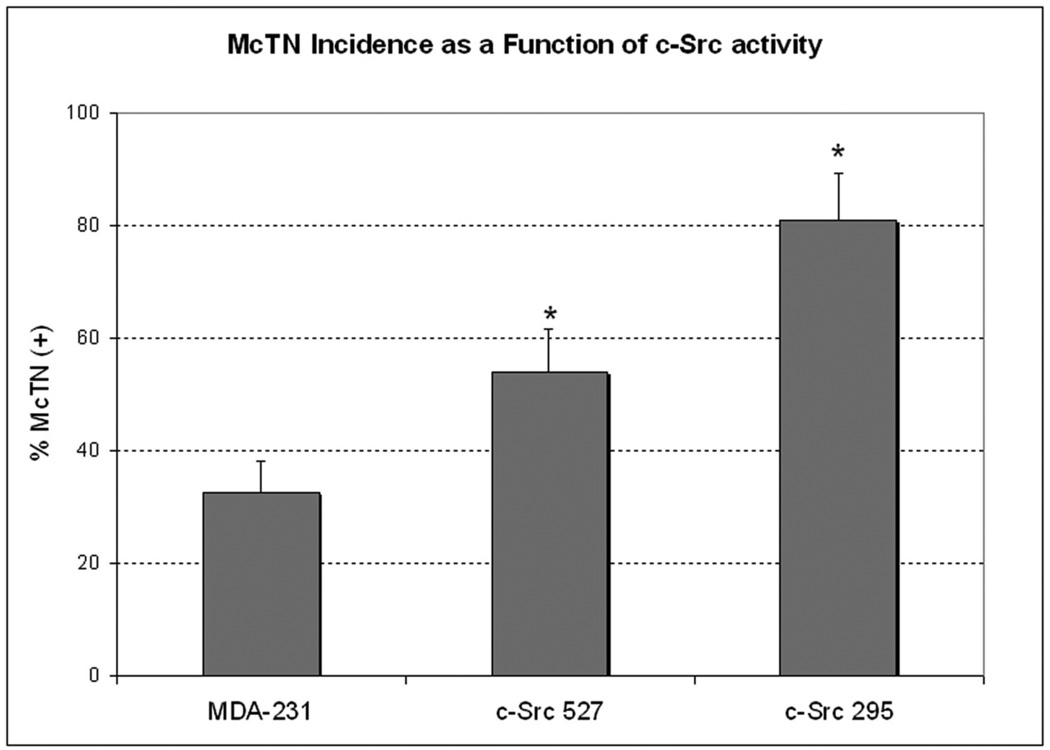
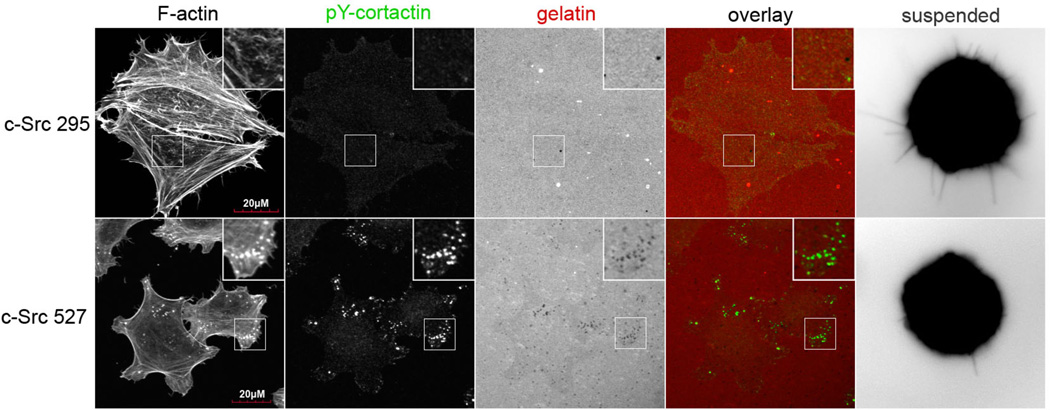
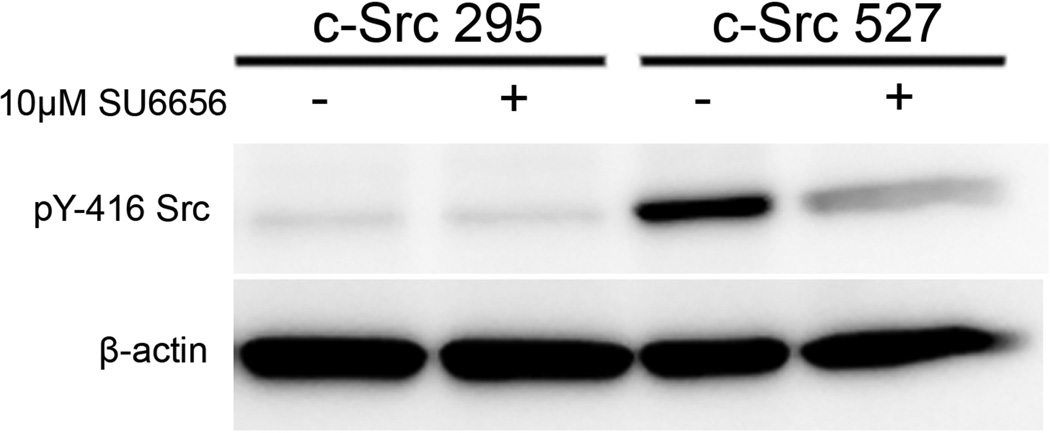
During metastasis, invading cells produce various actin-based membrane protrusions that promote directional migration and proteolysis of extracellular matrix (ECM). Observations of actin staining within thin, tubulin-based microtentacle (McTN) protrusions in suspended MDA-MB-231 tumor cells, prompted an investigation of whether McTNs are structural or functional analogs of invadopodia. We show here that MDA-MB-231 cells are capable of producing invadopodia and McTNs, both of which contain F-actin. Invadopodium formation was enhanced by the expression of a constitutively active c-Src kinase, and repressed by the expression of dominant-negative, catalytically inactive form of c-Src. In contrast, expression of inactive c-Src significantly increased McTN formation. Direct inhibition of c-Src with the SU6656 inhibitor compound also significantly enhanced McTN formation, but suppressed invadopodia, including the appearance of F-actin cores and phospho-cortactin foci, as well as completely blocking focal degradation of ECM. In addition, silencing of Tks5 in Src-transformed fibroblasts blocked invadopodia without affecting McTNs. Genetic modification of c-Src activity that promoted McTN formation augmented capillary retention of circulating tumor cells in vivo and rapid re-attachment of suspended cells in vitro, even though invadopodia were strongly suppressed. These results indicate that McTNs are capable of enhancing tumor cell reattachment, even in the absence of Tks5 and active Src, and define separate cytoskeletal mechanisms and functions for McTNs and invadopodia.
Conflict of interest statement
Figures
References
-
- Yilmaz M, Christofori G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009;28(1–2):15–33. - PubMed
-
- Gupton SL, Gertler FB. Filopodia: the fingers that do the walking. Sci STKE. 2007;2007(400):re5. - PubMed
-
- Machesky LM. Lamellipodia and filopodia in metastasis and invasion. FEBS Lett. 2008;582(14):2102–2111. - PubMed
-
- Linder S. The matrix corroded: podosomes and invadopodia in extracellular matrix degradation. Trends Cell Biol. 2007;17(3):107–117. - PubMed
-
- Buccione R, Caldieri G, Ayala I. Invadopodia: specialized tumor cell structures for the focal degradation of the extracellular matrix. Cancer Metastasis Rev. 2009;28(1–2):137–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
