

Methanol assimilation in Methylobacterium extorquens AM1: demonstration of all enzymes and their regulation
- PMID: 20957036
- PMCID: PMC2948502
- DOI: 10.1371/journal.pone.0013001
Methanol assimilation in Methylobacterium extorquens AM1: demonstration of all enzymes and their regulation
Abstract
Background: Methylobacterium extorquens AM1 is an aerobic facultative methylotrophic α-proteobacterium that can use reduced one-carbon compounds such as methanol, but also multi-carbon substrates like acetate (C₂) or succinate (C₄) as sole carbon and energy source. The organism has gained interest as future biotechnological production platform based on methanol as feedstock.
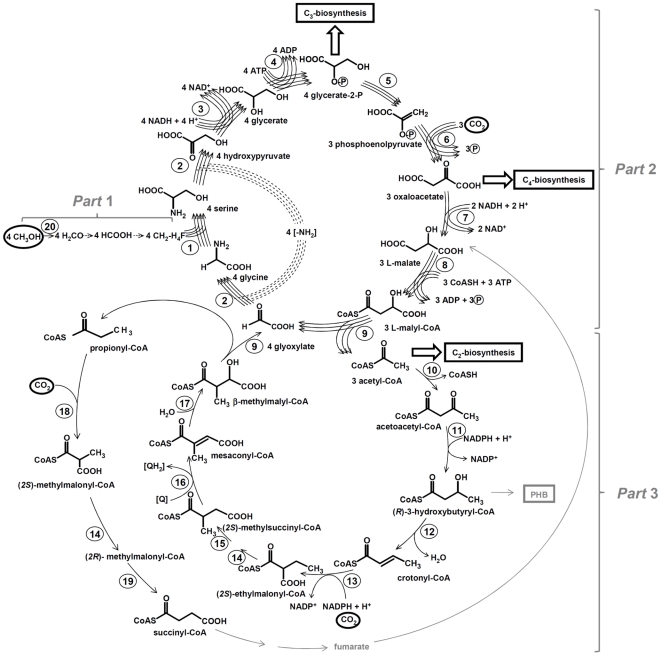
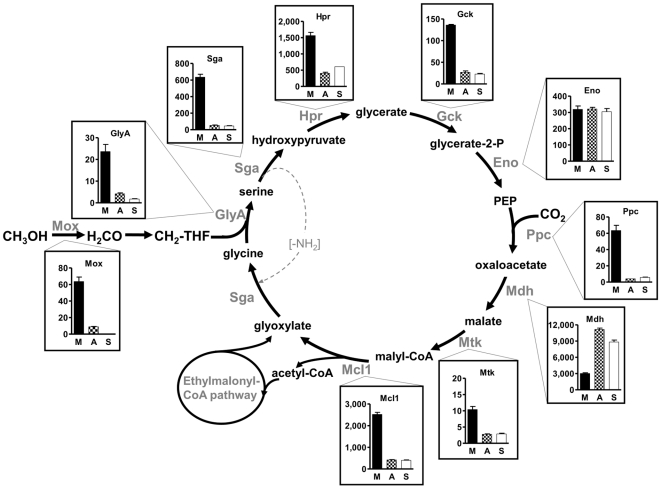
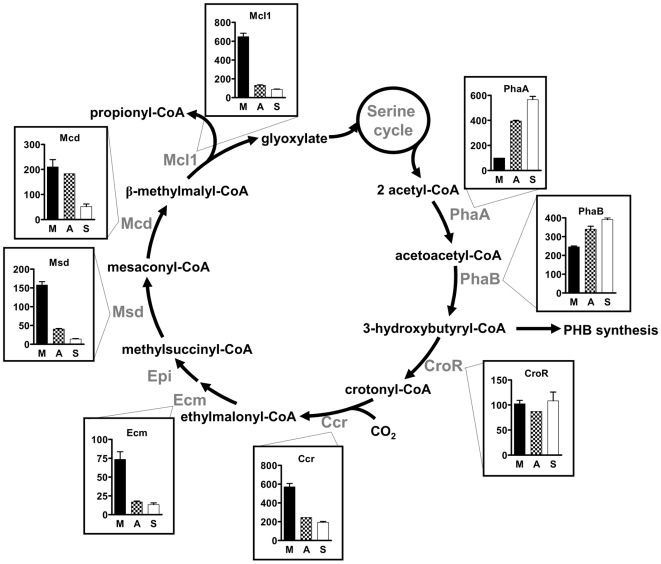
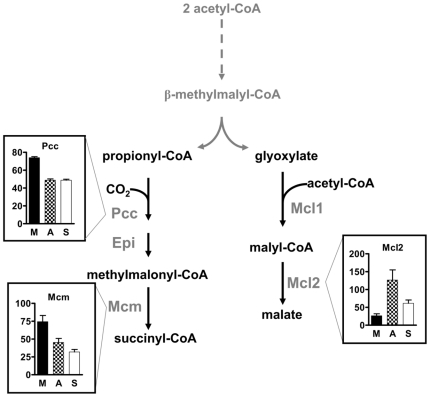
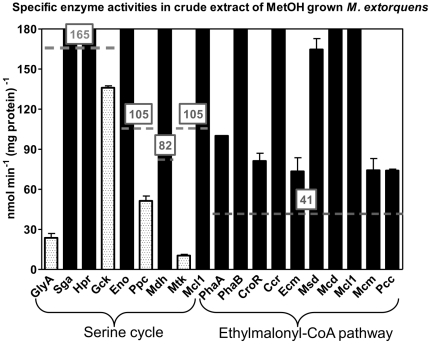
Methodology/principal findings: We present a comprehensive study of all postulated enzymes for the assimilation of methanol and their regulation in response to the carbon source. Formaldehyde, which is derived from methanol oxidation, is assimilated via the serine cycle, which starts with glyoxylate and forms acetyl-CoA. Acetyl-CoA is assimilated via the proposed ethylmalonyl-CoA pathway, which thereby regenerates glyoxylate. To further the understanding of the central carbon metabolism we identified and quantified all enzymes of the pathways involved in methanol assimilation. We observed a strict differential regulation of their activity level depending on whether C₁, C₂ or C₄ compounds are used. The enzymes, which are specifically required for the utilization of the individual substrates, were several-fold up-regulated and those not required were down-regulated. The enzymes of the ethylmalonyl-CoA pathway showed specific activities, which were higher than the calculated minimal values that can account for the observed growth rate. Yet, some enzymes of the serine cycle, notably its first and last enzymes serine hydroxymethyl transferase and malate thiokinase, exhibit much lower values and probably are rate limiting during methylotrophic growth. We identified the natural C₁ carrying coenzyme as tetrahydropteroyl-tetraglutamate rather than tetrahydrofolate.
Conclusion/significance: This study provides the first complete picture of the enzymes required for methanol assimilation, the regulation of their activity levels in response to the growth substrate, and the identification of potential growth limiting steps.
Conflict of interest statement
Figures
Similar articles
-
Replacing the Ethylmalonyl-CoA Pathway with the Glyoxylate Shunt Provides Metabolic Flexibility in the Central Carbon Metabolism of Methylobacterium extorquens AM1.ACS Synth Biol. 2018 Jan 19;7(1):86-97. doi: 10.1021/acssynbio.7b00229. Epub 2017 Dec 21. ACS Synth Biol. 2018. PMID: 29216425
-
Production of 3-hydroxypropionic acid in engineered Methylobacterium extorquens AM1 and its reassimilation through a reductive route.Microb Cell Fact. 2017 Oct 30;16(1):179. doi: 10.1186/s12934-017-0798-2. Microb Cell Fact. 2017. PMID: 29084554 Free PMC article.
-
Ethylmalonyl coenzyme A mutase operates as a metabolic control point in Methylobacterium extorquens AM1.J Bacteriol. 2015 Feb 15;197(4):727-35. doi: 10.1128/JB.02478-14. Epub 2014 Dec 1. J Bacteriol. 2015. PMID: 25448820 Free PMC article.
-
Biotechnological potential of the ethylmalonyl-CoA pathway.Appl Microbiol Biotechnol. 2011 Jan;89(1):17-25. doi: 10.1007/s00253-010-2873-z. Epub 2010 Sep 30. Appl Microbiol Biotechnol. 2011. PMID: 20882276 Review.
-
Methylobacterium extorquens: methylotrophy and biotechnological applications.Appl Microbiol Biotechnol. 2015 Jan;99(2):517-34. doi: 10.1007/s00253-014-6240-3. Epub 2014 Nov 30. Appl Microbiol Biotechnol. 2015. PMID: 25432674 Review.
Cited by
-
Carboxylases in natural and synthetic microbial pathways.Appl Environ Microbiol. 2011 Dec;77(24):8466-77. doi: 10.1128/AEM.05702-11. Epub 2011 Oct 14. Appl Environ Microbiol. 2011. PMID: 22003013 Free PMC article. Review.
-
Sedimentimonas flavescens gen. nov., sp. nov., isolated from sediment of Clam Island, Liaoning Province.Antonie Van Leeuwenhoek. 2022 Aug;115(8):979-994. doi: 10.1007/s10482-022-01754-4. Epub 2022 Jun 7. Antonie Van Leeuwenhoek. 2022. PMID: 35672593
-
Impact of glyphosate and glyphosate-based herbicides on phyllospheric Methylobacterium.BMC Plant Biol. 2024 Feb 19;24(1):119. doi: 10.1186/s12870-024-04818-x. BMC Plant Biol. 2024. PMID: 38369476 Free PMC article.
-
A survey of Methylobacterium species and strains reveals widespread production and varying profiles of cytokinin phytohormones.BMC Microbiol. 2022 Feb 8;22(1):49. doi: 10.1186/s12866-022-02454-9. BMC Microbiol. 2022. PMID: 35135483 Free PMC article.
-
Extensive Thioautotrophic Gill Endosymbiont Diversity within a Single Ctena orbiculata (Bivalvia: Lucinidae) Population and Implications for Defining Host-Symbiont Specificity and Species Recognition.mSystems. 2019 Aug 27;4(4):e00280-19. doi: 10.1128/mSystems.00280-19. mSystems. 2019. PMID: 31455638 Free PMC article.
References
-
- Madhaiyan M, Poonguzhali S, Senthilkumar M, Seshadri S, Chung J, et al. Growth promotion and induction of systemic resistance in rice cultivar Co-47 (Oryza sativa L.) by Methylobacterium spp. Bot Bull Acad Sin. 2004;45(4):315–324.
-
- Kirstine WV, Galbally IE. A simple model for estimating emissions of volatile organic compounds from grass and cut grass in urban airsheds and its application to two Australian cities. J Air & Waste Manag Assoc. 2004;54:1299–1311. - PubMed
-
- Abanda-Nkpwatt D, Müsch M, Tschiersch J, Boettner M, Schwab W. Molecular interaction between Methylobacterium extorquens and seedlings: growth promotion, methanol consumption, and localization of the methanol emission site. J Experimental Botany. 2006;57(15):4025–4032. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
