

CD300 heterocomplexes, a new and family-restricted mechanism for myeloid cell signaling regulation
- PMID: 20959446
- PMCID: PMC3009906
- DOI: 10.1074/jbc.M110.140889
CD300 heterocomplexes, a new and family-restricted mechanism for myeloid cell signaling regulation
Abstract
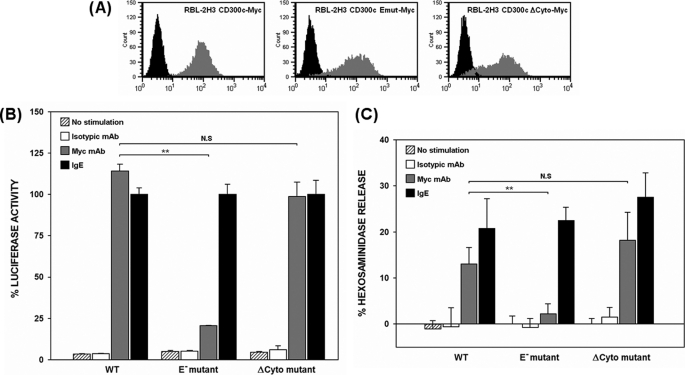
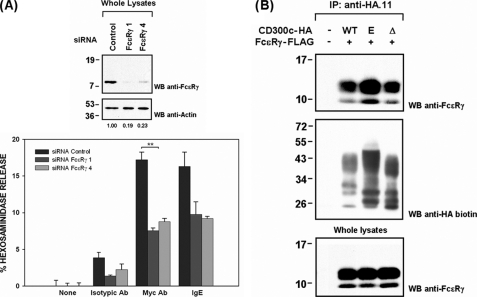
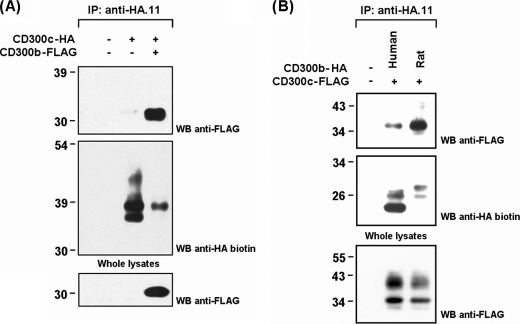
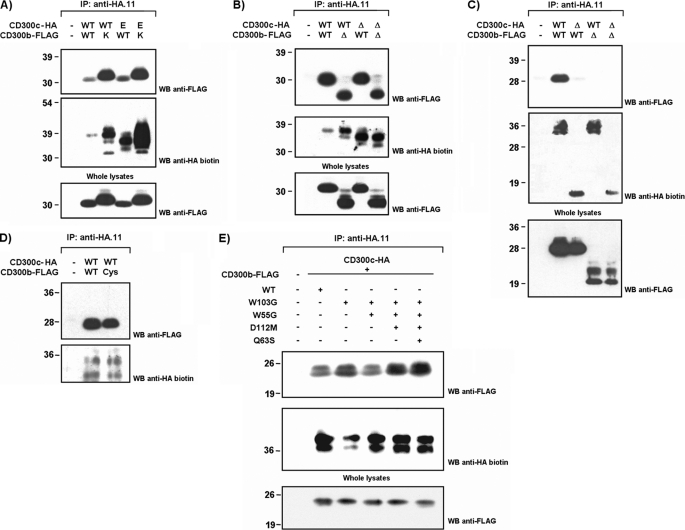
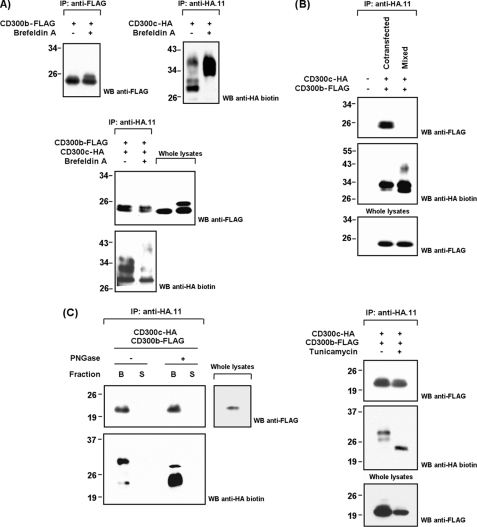
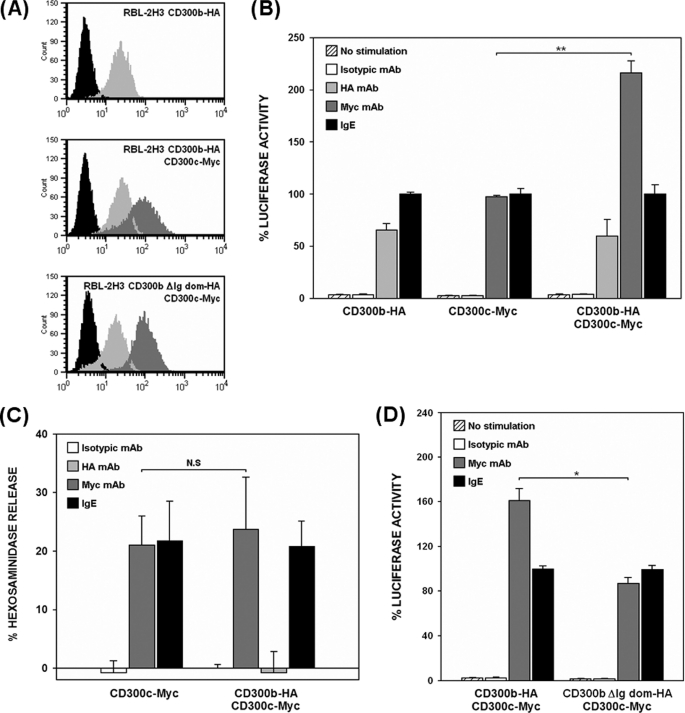
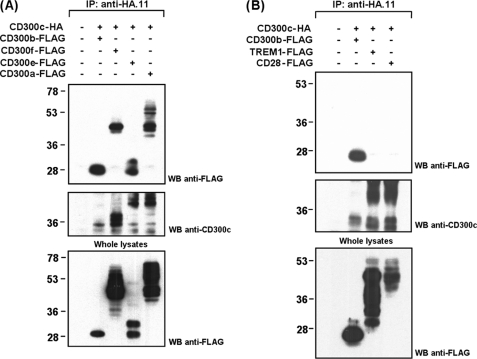
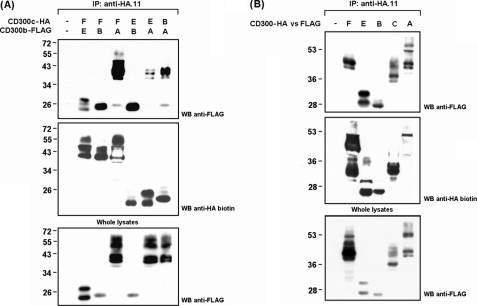
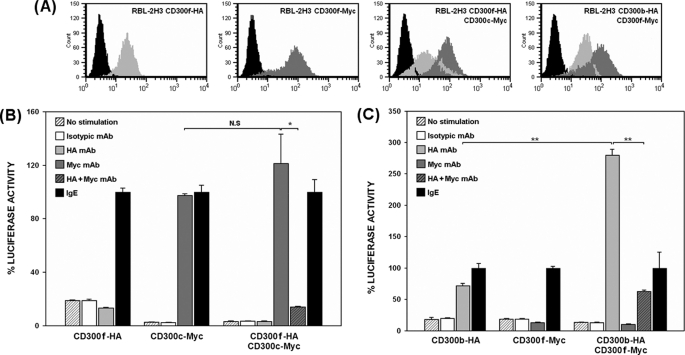
The CD300 family of myeloid immunoglobulin receptors includes activating (CD300b, CD300e) and inhibitory members (CD300a, CD300f), as well as molecules of uncertain function presenting a negative charge within their transmembrane domain (CD300c, CD300d). In this paper, we establish that CD300c is a functional immune receptor able to deliver activating signals upon ligation in RBL-2H3 mast cells. CD300c signaling is partially mediated by a direct association with the immune receptor tyrosine-based activation motif-bearing adaptor FcεRγ. The existence of complementary transmembrane-charged residues in certain CD300 receptors suggested the formation of heterodimers within this family. Indeed, we proved the interaction between CD300b and CD300c in transfected COS-7 cells and demonstrated that it has important functional consequences. Unexpectedly, dimmer formation was dependent on the immunoglobulin domains rather than the charged transmembrane residues. Concordantly, all CD300 members were found to interact with each other, even with themselves, forming both homo- and heterodimers. We found that the combination of CD300 receptors in a complex differentially modulates the signaling outcome, strongly suggesting a new mechanism by which CD300 complexes could regulate the activation of myeloid cells upon interaction with their natural ligands.
Figures
References
-
- Veillette A., Latour S., Davidson D. (2002) Annu. Rev. Immunol. 20, 669–707 - PubMed
-
- Tomasello E., Bléry M., Vély F., Vivier E. (2000) Semin. Immunol. 12, 139–147 - PubMed
-
- Humphrey M. B., Lanier L. L., Nakamura M. C. (2005) Immunol. Rev. 208, 50–65 - PubMed
-
- Underhill D. M., Goodridge H. S. (2007) Trends Immunol. 28, 66–73 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
