

Inhibition of permeability transition pore opening by mitochondrial STAT3 and its role in myocardial ischemia/reperfusion
- PMID: 20960209
- PMCID: PMC2978889
- DOI: 10.1007/s00395-010-0124-1
Inhibition of permeability transition pore opening by mitochondrial STAT3 and its role in myocardial ischemia/reperfusion
Abstract
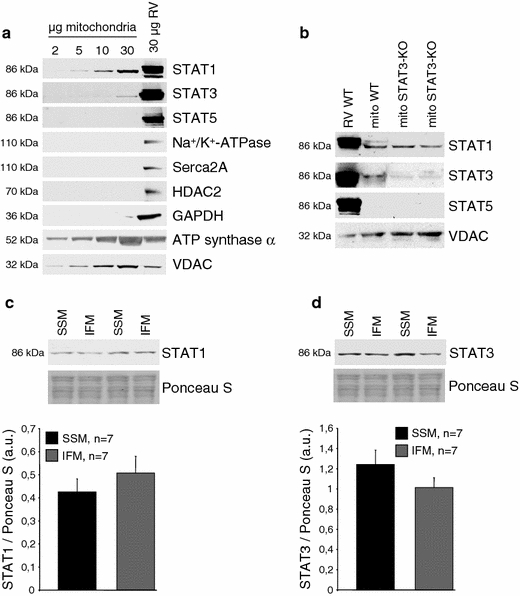
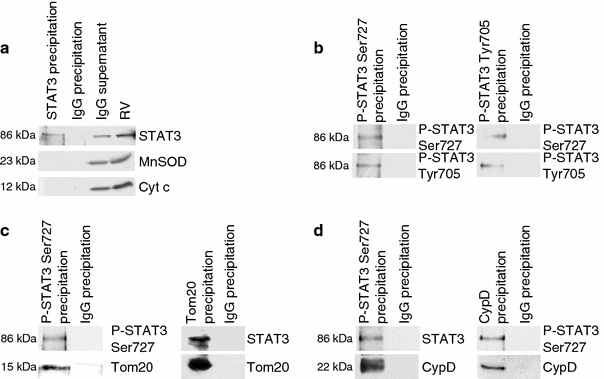
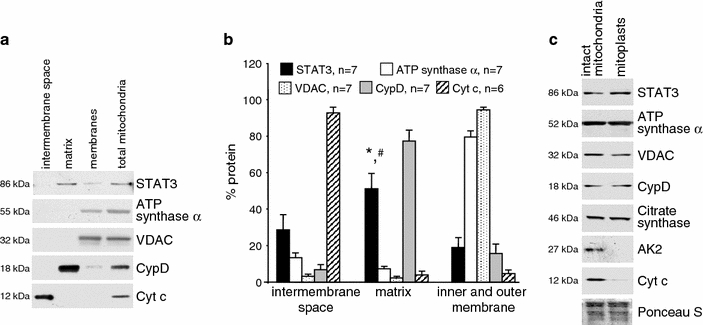
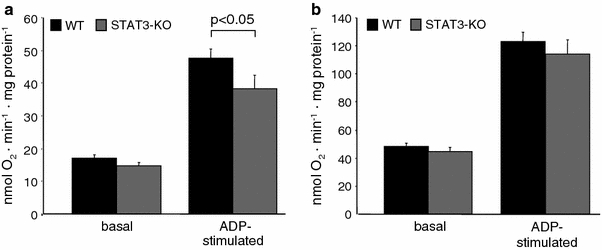
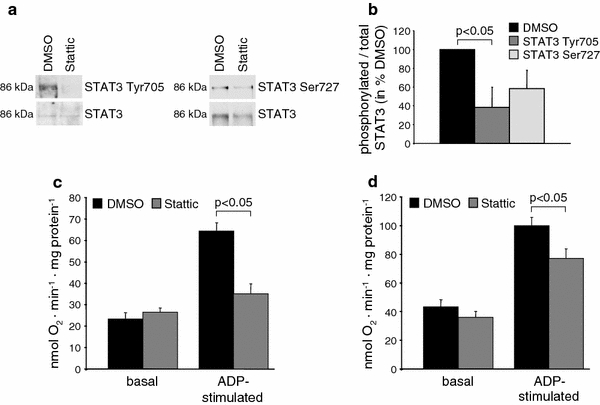
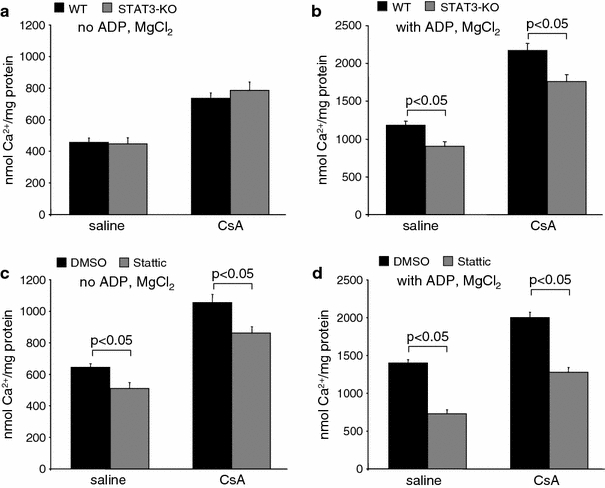
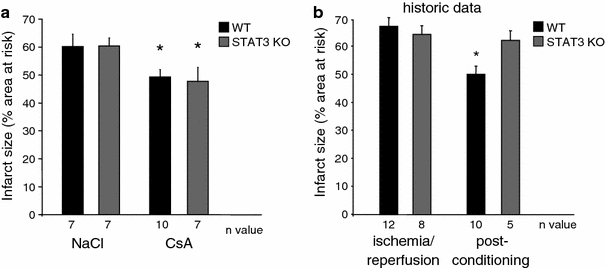
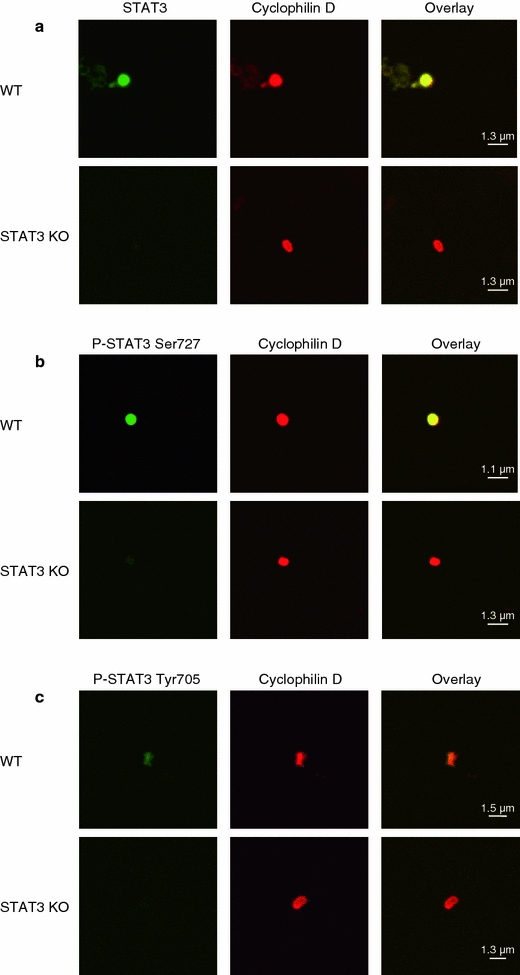
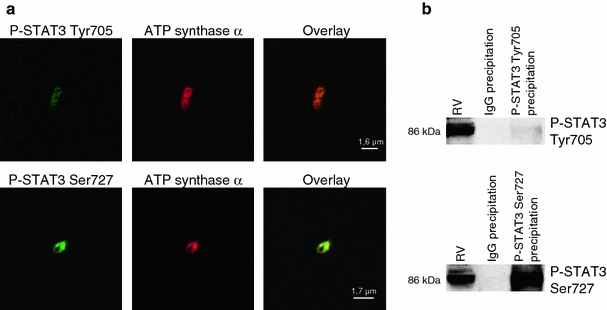
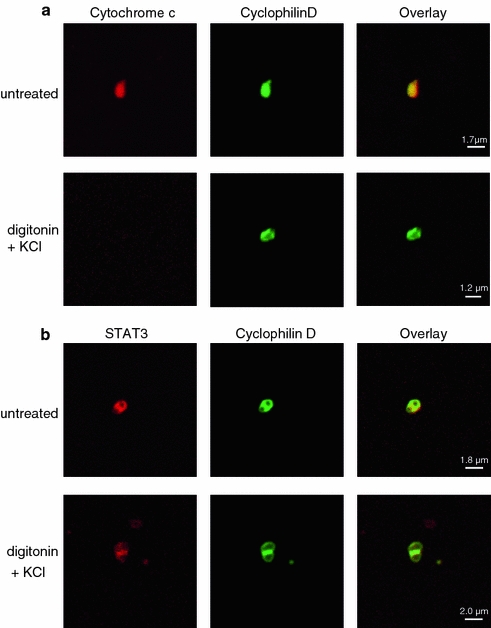
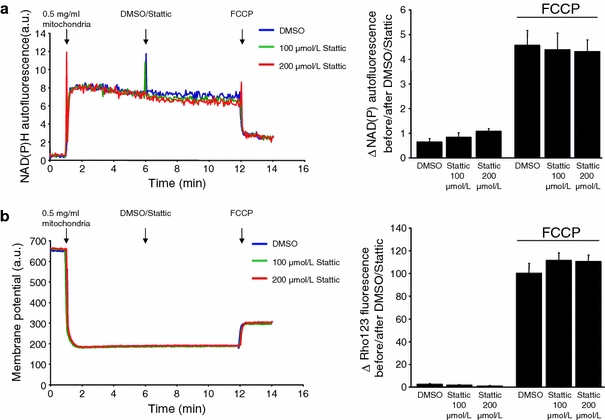
The signal transducer and activator of transcription 3 (STAT3) contributes to cardioprotection by ischemic pre- and postconditioning. Mitochondria are central elements of cardioprotective signaling, most likely by delaying mitochondrial permeability transition pore (MPTP) opening, and STAT3 has recently been identified in mitochondria. We now characterized the mitochondrial localization of STAT3 and its impact on respiration and MPTP opening. STAT3 was mainly present in the matrix of subsarcolemmal and interfibrillar cardiomyocyte mitochondria. STAT1, but not STAT5 was also detected in mitochondria under physiological conditions. ADP-stimulated respiration was reduced in mitochondria from mice with a cardiomyocyte-specific deletion of STAT3 (STAT3-KO) versus wildtypes and in rat mitochondria treated with the STAT3 inhibitor Stattic (STAT3 inhibitory compound, 6-Nitrobenzo[b]thiophene 1,1-dioxide). Mitochondria from STAT3-KO mice and Stattic-treated rat mitochondria tolerated less calcium until MPTP opening occurred. STAT3 co-immunoprecipitated with cyclophilin D, the target of the cardioprotective agent and MPTP inhibitor cyclosporine A (CsA). However, CsA reduced infarct size to a similar extent in wildtype and STAT3-KO mice in vivo. Thus, STAT3 possibly contributes to cardioprotection by stimulation of respiration and inhibition of MPTP opening.
Figures
References
-
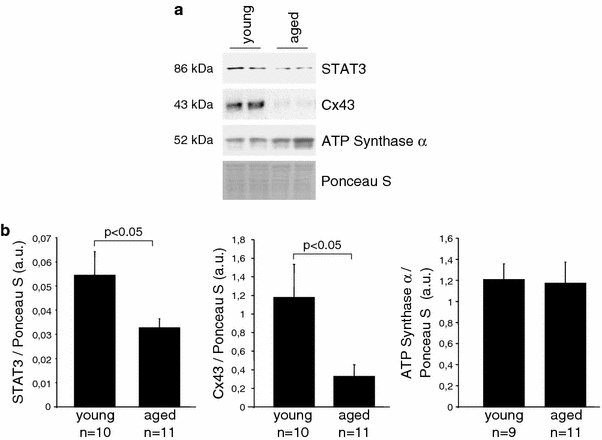
- Boengler K, Dodoni G, Rodriguez-Sinovas A, Cabestrero A, Ruiz-Maena M, Gres P, Konietzka I, Lopez-Iglesias C, García-Dorado D, Heusch G, Schulz R. Connexin 43 in cardiomyocyte mitochondria and its increase by ischemic preconditioning. Cardiovasc Res. 2005;67:234–244. doi: 10.1016/j.cardiores.2005.04.014. - DOI - PubMed
-
- Boengler K, Konietzka I, Buechert A, Heinen Y, Garcia-Dorado D, Heusch G, Schulz R. Loss of ischemic preconditioning’s cardioprotection in aged mouse hearts is associated with reduced gap junctional and mitochondrial levels of connexin 43. Am J Physiol Heart Circ Physiol. 2007;292:H1764–H1769. doi: 10.1152/ajpheart.01071.2006. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
