

Lessons from genome-wide studies: an integrated definition of the coactivator function of histone acetyl transferases
- PMID: 20961410
- PMCID: PMC2972259
- DOI: 10.1186/1756-8935-3-18
Lessons from genome-wide studies: an integrated definition of the coactivator function of histone acetyl transferases
Abstract
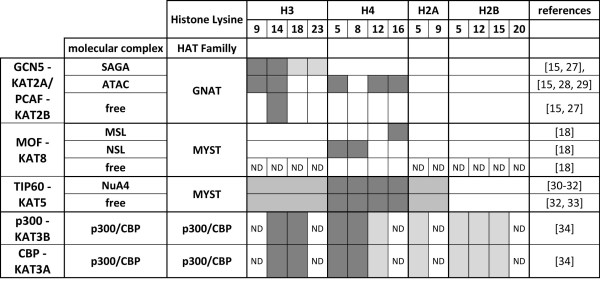
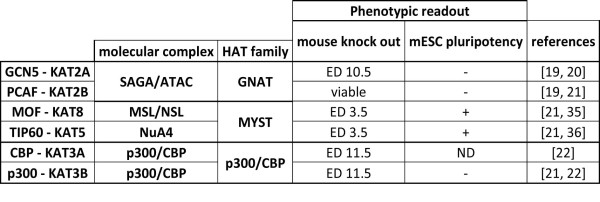
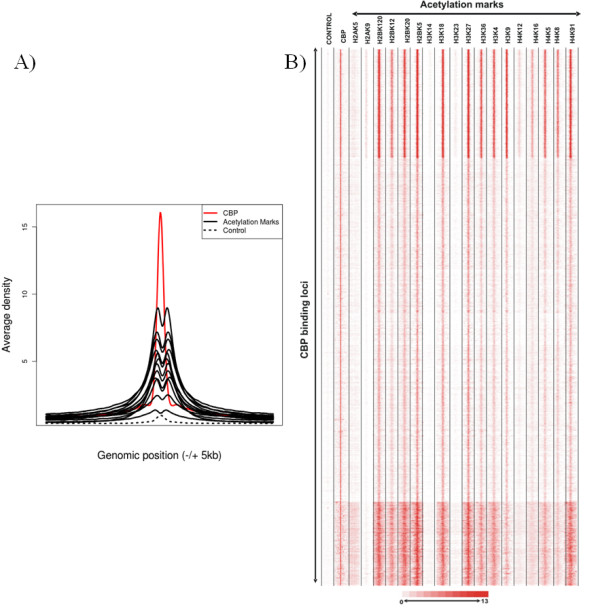
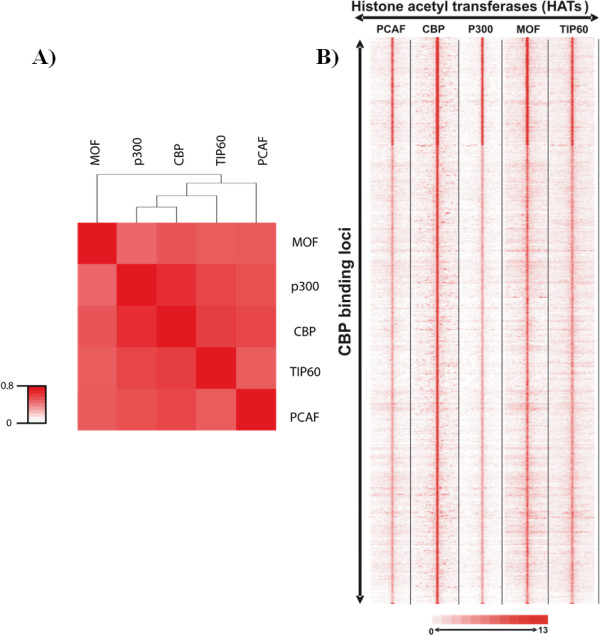
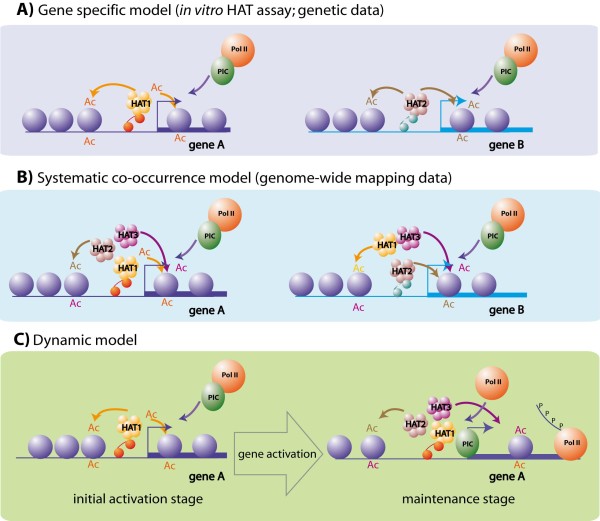
Histone acetylation is one of the key regulatory mechanisms controlling transcriptional activity in eukaryotic cells. In higher eukaryotes, a number of nuclear histone acetyltransferase (HAT) enzymes have been identified, most of which are part of a large multisubunit complex. This diversity, combined with the large number of potentially acetylable lysines on histones, suggested the existence of a specific regulatory mechanism based on the substrate specificity of HATs. Over the past decade, intensive characterisations of the HAT complexes have been carried out. However, the precise mode of action of HATs, and particularly the functional differences amongst these complexes, remains elusive. Here we review current insights into the functional role of HATs, focusing on the specificity of their action. Studies based on biochemical as well as genetic approaches suggested that HATs exert a high degree of specificity in their acetylation spectra and in the cellular processes they regulate. However, a different view emerged recently from genomic approaches that provided genome-wide maps of HAT recruitments. The careful analysis of genomic data suggests that all HAT complexes would be simultaneously recruited to a similar set of loci in the genome, arguing for a low specificity in their function. In this review, we discuss the significance of these apparent contradictions and suggest a new model that integrates biochemical, genetic and genome-wide data to better describe the functional specificity of HAT complexes.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous
