

doi: 10.1101/gad.1989510.
Epub 2010 Oct 21.
Roles for the Yb body components Armitage and Yb in primary piRNA biogenesis in Drosophila
Affiliations
- PMID: 20966047
- PMCID: PMC2975925
- DOI: 10.1101/gad.1989510
Item in Clipboard
Roles for the Yb body components Armitage and Yb in primary piRNA biogenesis in Drosophila
Genes Dev.
.
Abstract
PIWI-interacting RNAs (piRNAs) protect genome integrity from transposons. In Drosophila ovarian somas, primary piRNAs are produced and loaded onto Piwi. Here, we describe roles for the cytoplasmic Yb body components Armitage and Yb in somatic primary piRNA biogenesis. Armitage binds to Piwi and is required for localizing Piwi into Yb bodies. Without Armitage or Yb, Piwi is freed from the piRNAs and does not enter the nucleus. Thus, piRNA loading is required for Piwi nuclear entry. We propose that a functional Piwi-piRNA complex is formed and inspected in Yb bodies before its nuclear entry to exert transposon silencing.
Figures
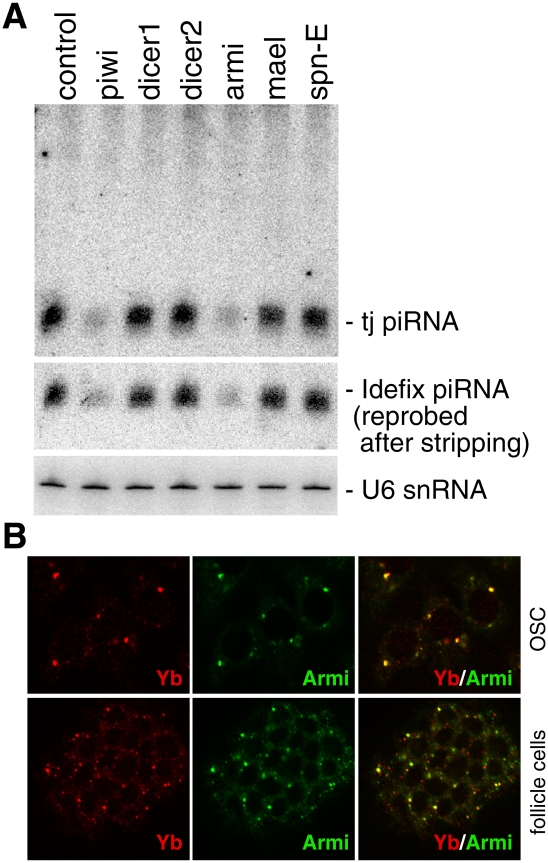
Armi, a novel component of Yb bodies, is necessary for primary piRNA production. (A) RNAi-based depletion of Armi, but not of Spn-E and Mael, caused a severe reduction in the expression levels of tj-piRNA and Idefix-piRNA in OSCs. (B) OSCs and ovaries immunostained with anti-Armi and anti-Yb antibodies. Merged images show that Armi colocalizes with Yb at Yb bodies.
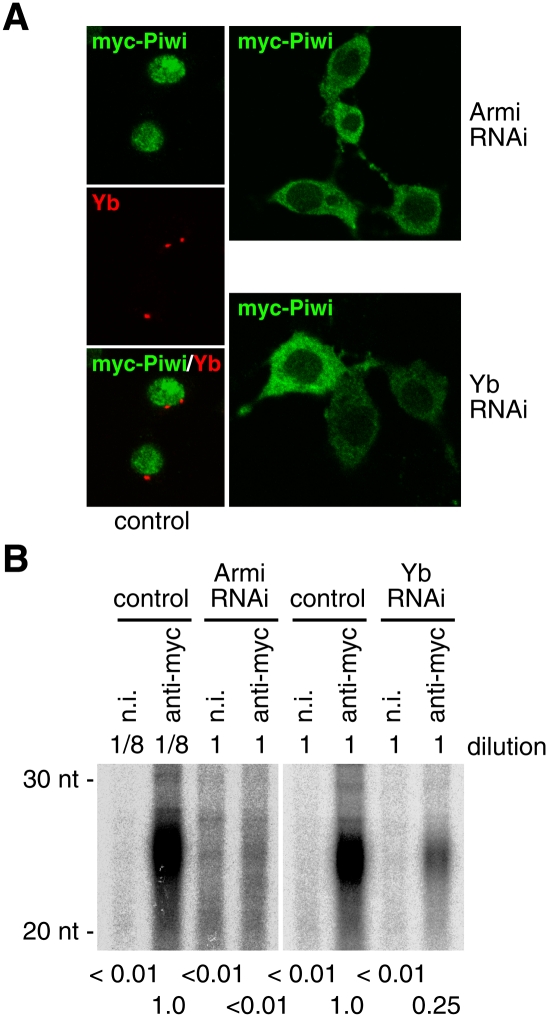
Depletion of Armi and Yb causes Piwi to be mislocalized to the cytoplasm. (A) After depletion of Armi and Yb, myc-tagged Piwi was expressed in OSCs by transfection. myc-Piwi accumulated predominantly in the cytoplasm, suggesting that the nuclear signals observed in Supplemental Figure S3B (green) reflect endogenous Piwi that was localized to the nucleus prior to Armi and Yb depletion. (B) 32P-labeling of RNAs showed that, when Armi or Yb was depleted, myc-Piwi was loaded with few or no piRNAs. (Left panel) Note that the complex immunoprecipitated from control cells was diluted 1:8 prior to RNA isolation. The amount of Piwi in the immunoprecipitated complex from control cells was eightfold higher than that from Armi RNAi cells (data not shown).
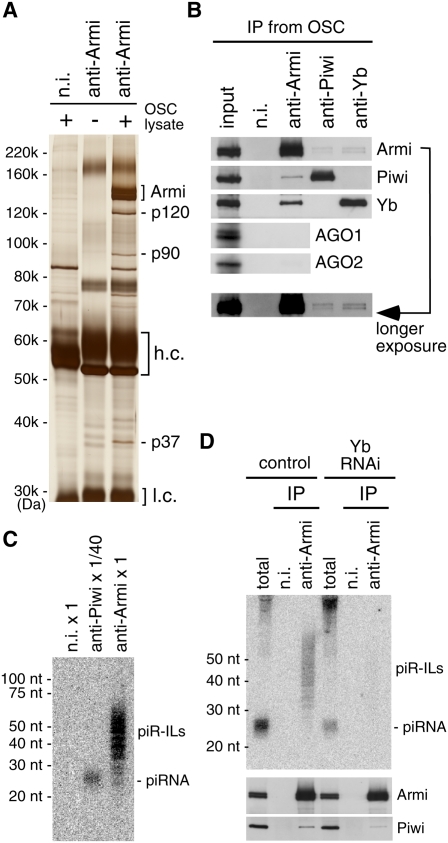
The Armi complex contains not only Piwi and Yb, but also piR-ILs. (A) Silver staining of protein components in the immunoprecipitated complex with anti-Armi antibody from OSCs. Three proteins—p120, p90, and p37—coimmunoprecipitated with Armi. (B) The complex immunoprecipitated with anti-Armi antibody contains Piwi and Yb but not AGO1 and AGO2. Yb was not detected in the complex. In addition, Piwi was not detected in the complex immunoprecipitated by anti-Yb. The input lane contains 1% of lysates used for immunoprecipitation. (C) The Armi complex immunopurified from OSCs contains piR-ILs. RNA molecules isolated from the complexes immunoprecipitated with anti-Piwi and anti-Armi antibodies were probed with a DNA oligonucleotide recognizing tj-piRNA. Note that the Piwi complex was diluted 1:40 prior to RNA isolation. (D, top panel) Depletion of Yb resulted in the disappearance of piR-ILs from the Armi complex. (Bottom panels) Western blotting showed that the association between Piwi and Armi was maintained under these conditions.
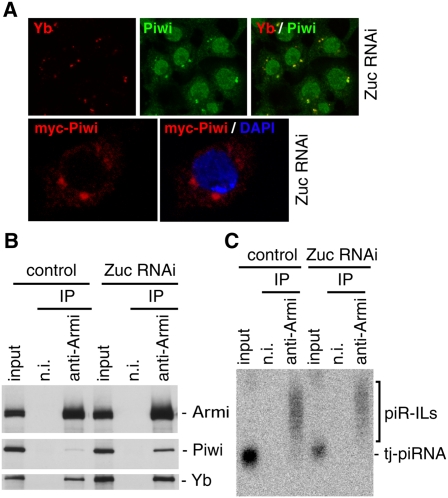
Zuc functions in the primary piRNA processing pathway in OSCs. (A, top panels) Depletion of Zuc caused Piwi (green) to be localized to Yb bodies. Yb localization (red) was not affected by Zuc depletion. (Bottom panels) myc-Piwi (red) was expressed by transfection after Zuc was depleted in OSCs by RNAi. (B) Depletion of Zuc did not affect the association of Armi with Piwi and Yb. Under such conditions, Piwi was slightly more abundant in the Armi–Piwi–Yb complex. (C) Depletion of Zuc did not affect piRNA intermediates in the Armi complex.
References
-
- Aravin AA, Hannon GJ, Brennecke J 2007. The Piwi–piRNA pathway provides an adaptive defense in the transposon arms race. Science 318: 761–764 - PubMed
-
- Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ 2007. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128: 1089–1103 - PubMed
-
- Choi SY, Huang P, Jenkins GM, Chan DC, Schiller J, Frohman MA 2006. A common lipid links Mfn-mediated mitochondrial fusion and SNARE-regulated exocytosis. Nat Cell Biol 8: 1255–1262 - PubMed
-
- Cook HA, Koppetsch BS, Wu J, Theurkauf WE 2004. The Drosophila SDE3 homolog armitage is required for oskar mRNA silencing and embryonic axis specification. Cell 116: 817–829 - PubMed
-
- Cox DN, Chao A, Lin H 2000. piwi encodes a nucleoplasmic factor whose activity modulates the number and division rate of germline stem cells. Development 127: 503–514 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials