

Phosphorylation of p50 NF-kappaB at a single serine residue by DNA-dependent protein kinase is critical for VCAM-1 expression upon TNF treatment
- PMID: 20966071
- PMCID: PMC3003413
- DOI: 10.1074/jbc.M110.158352
Phosphorylation of p50 NF-kappaB at a single serine residue by DNA-dependent protein kinase is critical for VCAM-1 expression upon TNF treatment
Abstract
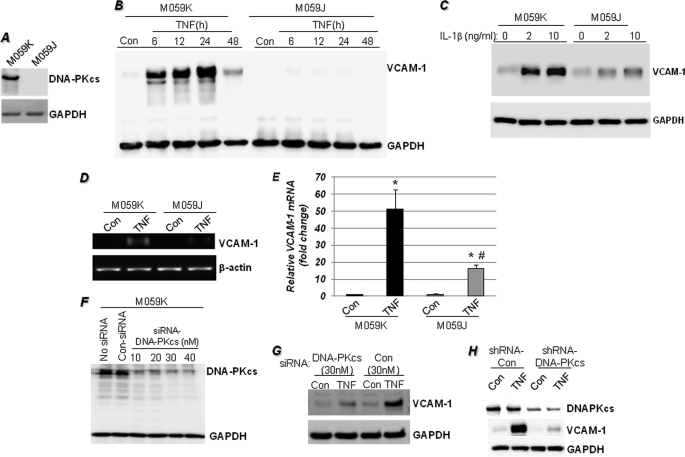
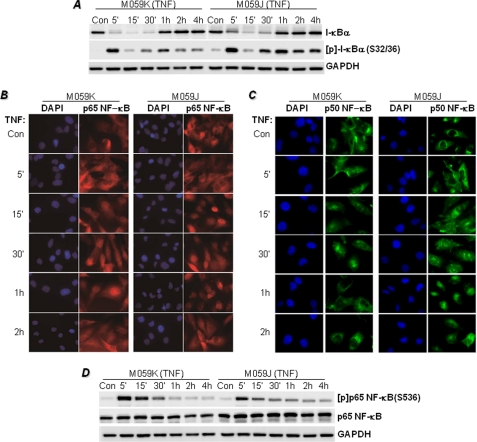
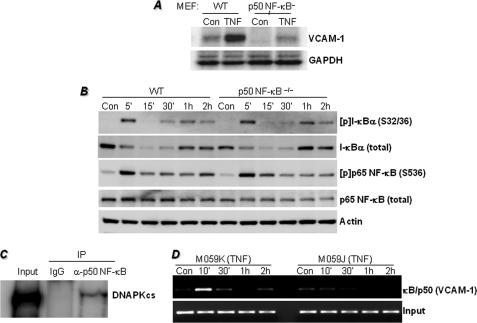
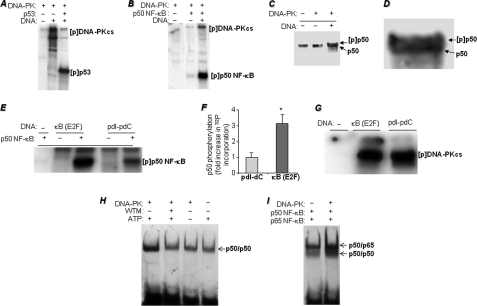
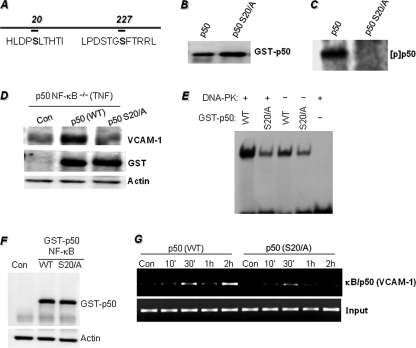
The DNA binding activity of NF-κB is critical for VCAM-1 expression during inflammation. DNA-dependent protein kinase (DNA-PK) is thought to be involved in NF-κB activation. Here we show that DNA-PK is required for VCAM-1 expression in response to TNF. The phosphorylation and subsequent degradation of I-κBα as well as the serine 536 phosphorylation and nuclear translocation of p65 NF-κB were insufficient for VCAM-1 expression in response to TNF. The requirement for p50 NF-κB in TNF-induced VCAM-1 expression may be associated with its interaction with and phosphorylation by DNA-PK, which appears to be dominant over the requirement for p65 NF-κB activation. p50 NF-κB binding to its consensus sequence increased its susceptibility to phosphorylation by DNA-PK. Additionally, DNA-PK activity appeared to increase the association between p50/p50 and p50/p65 NF-κB dimers upon binding to DNA and after binding of p50 NF-κB to the VCAM-1 promoter. Analyses of the p50 NF-κB protein sequence revealed that both serine 20 and serine 227 at the amino terminus of the protein are putative sites for phosphorylation by DNA-PK. Mutation of serine 20 completely eliminated phosphorylation of p50 NF-κB by DNA-PK, suggesting that serine 20 is the only site in p50 NF-κB for phosphorylation by DNA-PK. Re-establishing wild-type p50 NF-κB, but not its serine 20/alanine mutant, in p50 NF-κB(-/-) fibroblasts reversed VCAM-1 expression after TNF treatment, demonstrating the importance of the serine 20 phosphorylation site in the induction of VCAM-1 expression. Together, these results elucidate a novel mechanism for the involvement of DNA-PK in the positive regulation of p50 NF-κB to drive VCAM-1 expression.
Figures
Similar articles
-
NF-kappa B regulates VCAM-1 expression on fibroblast-like synoviocytes.J Immunol. 2000 Jun 1;164(11):5990-7. doi: 10.4049/jimmunol.164.11.5990. J Immunol. 2000. PMID: 10820282
-
Requirement for both JAK-mediated PI3K signaling and ACT1/TRAF6/TAK1-dependent NF-kappaB activation by IL-17A in enhancing cytokine expression in human airway epithelial cells.J Immunol. 2007 Nov 15;179(10):6504-13. doi: 10.4049/jimmunol.179.10.6504. J Immunol. 2007. PMID: 17982039
-
Nuclear factor κB mediates suppression of canonical transient receptor potential 6 expression by reactive oxygen species and protein kinase C in kidney cells.J Biol Chem. 2013 May 3;288(18):12852-65. doi: 10.1074/jbc.M112.410357. Epub 2013 Mar 22. J Biol Chem. 2013. PMID: 23525112 Free PMC article.
-
Involvement of MAPKs and NF-kappaB in tumor necrosis factor alpha-induced vascular cell adhesion molecule 1 expression in human rheumatoid arthritis synovial fibroblasts.Arthritis Rheum. 2010 Jan;62(1):105-16. doi: 10.1002/art.25060. Arthritis Rheum. 2010. PMID: 20039412
-
The multifunctional role of intrinsic disorder in NF-κB signaling.Biochem Soc Trans. 2023 Dec 20;51(6):2085-2092. doi: 10.1042/BST20230035. Biochem Soc Trans. 2023. PMID: 38095058 Free PMC article. Review.
Cited by
-
Mechanistic link between DNA damage sensing, repairing and signaling factors and immune signaling.Adv Protein Chem Struct Biol. 2019;115:297-324. doi: 10.1016/bs.apcsb.2018.11.004. Epub 2019 Jan 3. Adv Protein Chem Struct Biol. 2019. PMID: 30798935 Free PMC article. Review.
-
DNA damage, metabolism and aging in pro-inflammatory T cells: Rheumatoid arthritis as a model system.Exp Gerontol. 2018 May;105:118-127. doi: 10.1016/j.exger.2017.10.027. Epub 2017 Nov 8. Exp Gerontol. 2018. PMID: 29101015 Free PMC article. Review.
-
NMR- and circular dichroism-monitored lipid binding studies suggest a general role for the FATC domain as membrane anchor of phosphatidylinositol 3-kinase-related kinases (PIKK).J Biol Chem. 2013 Jul 5;288(27):20046-63. doi: 10.1074/jbc.M113.467233. Epub 2013 May 13. J Biol Chem. 2013. PMID: 23671275 Free PMC article.
-
DNA-dependent protein kinase inhibition blocks asthma in mice and modulates human endothelial and CD4⁺ T-cell function without causing severe combined immunodeficiency.J Allergy Clin Immunol. 2015 Feb;135(2):425-40. doi: 10.1016/j.jaci.2014.09.005. Epub 2014 Oct 19. J Allergy Clin Immunol. 2015. PMID: 25441643 Free PMC article.
-
The regulation of sequence specific NF-κB DNA binding and transcription by IKKβ phosphorylation of NF-κB p50 at serine 80.Nucleic Acids Res. 2019 Dec 2;47(21):11151-11163. doi: 10.1093/nar/gkz873. Nucleic Acids Res. 2019. PMID: 31598684 Free PMC article.
References
-
- Braun M., Pietsch P., Schrör K., Baumann G., Felix S. B. (1999) Cardiovasc Res. 41, 395–401 - PubMed
-
- Makrilia N., Kollias A., Manolopoulos L., Syrigos K. (2009) Cancer Invest. 27, 1023–1037 - PubMed
-
- Rojas A. I., Ahmed A. R. (1999) Crit. Rev. Oral Biol. Med. 10, 337–358 - PubMed
-
- Huo Y., Ley K. (2001) Acta. Physiol. Scand. 173, 35–43 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
