

Towards a unifying, systems biology understanding of large-scale cellular death and destruction caused by poorly liganded iron: Parkinson's, Huntington's, Alzheimer's, prions, bactericides, chemical toxicology and others as examples
- PMID: 20967426
- PMCID: PMC2988997
- DOI: 10.1007/s00204-010-0577-x
Towards a unifying, systems biology understanding of large-scale cellular death and destruction caused by poorly liganded iron: Parkinson's, Huntington's, Alzheimer's, prions, bactericides, chemical toxicology and others as examples
Abstract
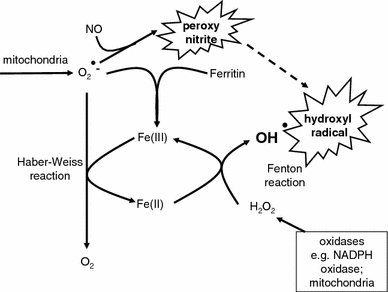
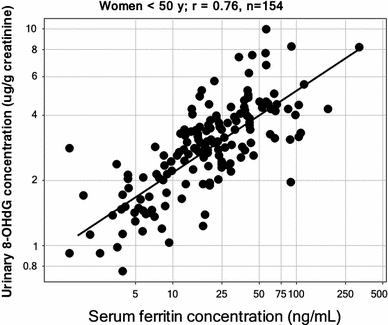
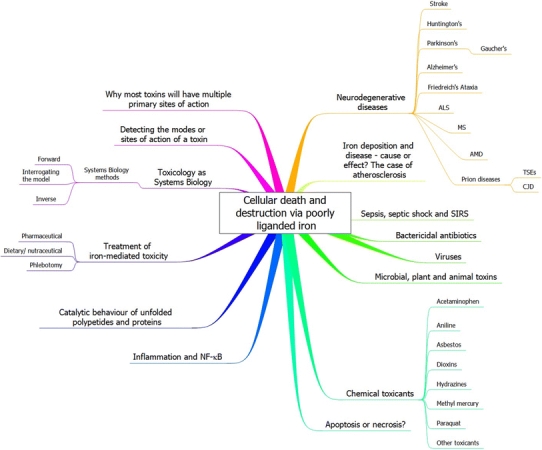
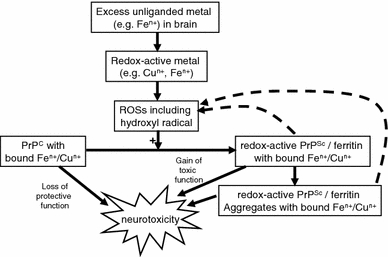
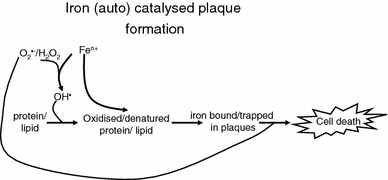
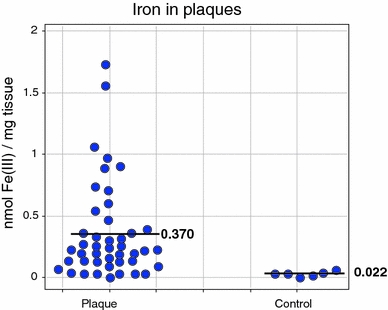
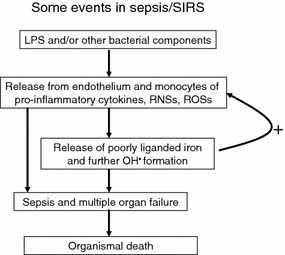
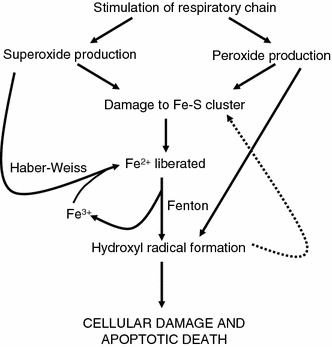
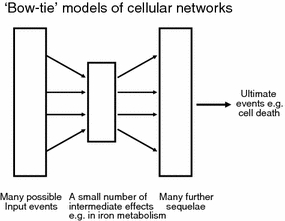
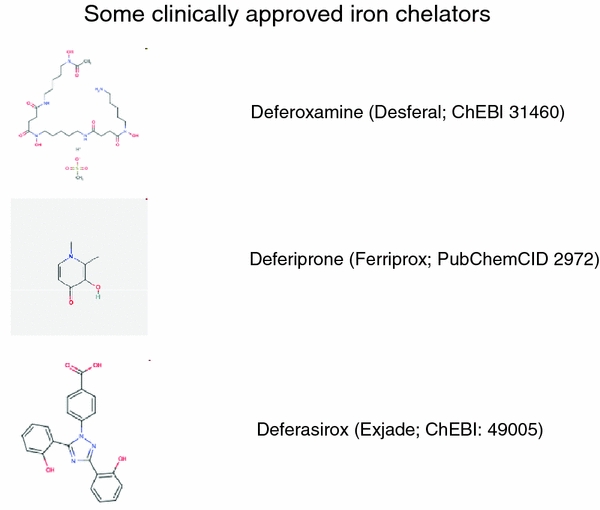
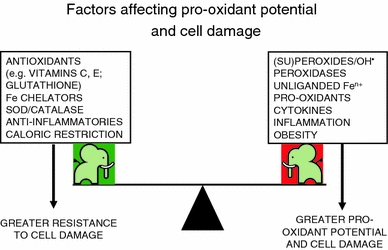
Exposure to a variety of toxins and/or infectious agents leads to disease, degeneration and death, often characterised by circumstances in which cells or tissues do not merely die and cease to function but may be more or less entirely obliterated. It is then legitimate to ask the question as to whether, despite the many kinds of agent involved, there may be at least some unifying mechanisms of such cell death and destruction. I summarise the evidence that in a great many cases, one underlying mechanism, providing major stresses of this type, entails continuing and autocatalytic production (based on positive feedback mechanisms) of hydroxyl radicals via Fenton chemistry involving poorly liganded iron, leading to cell death via apoptosis (probably including via pathways induced by changes in the NF-κB system). While every pathway is in some sense connected to every other one, I highlight the literature evidence suggesting that the degenerative effects of many diseases and toxicological insults converge on iron dysregulation. This highlights specifically the role of iron metabolism, and the detailed speciation of iron, in chemical and other toxicology, and has significant implications for the use of iron chelating substances (probably in partnership with appropriate anti-oxidants) as nutritional or therapeutic agents in inhibiting both the progression of these mainly degenerative diseases and the sequelae of both chronic and acute toxin exposure. The complexity of biochemical networks, especially those involving autocatalytic behaviour and positive feedbacks, means that multiple interventions (e.g. of iron chelators plus antioxidants) are likely to prove most effective. A variety of systems biology approaches, that I summarise, can predict both the mechanisms involved in these cell death pathways and the optimal sites of action for nutritional or pharmacological interventions.
Figures
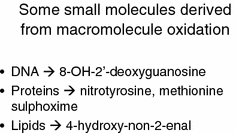

Comment in
-
Iron dysregulation: an important aspect in toxicology.Arch Toxicol. 2010 Nov;84(11):823-4. doi: 10.1007/s00204-010-0610-0. Arch Toxicol. 2010. PMID: 20941480 No abstract available.
References
-
- Abdelrahman M, Mazzon E, Bauer M, Bauer I, Delbosc S, Cristol JP, et al. Inhibitors of NADPH oxidase reduce the organ injury in hemorrhagic shock. Shock. 2005;23(2):107–114. - PubMed
-
- Abdelrahman M, Sharples EJ, McDonald MC, Collin M, Patel NS, Yaqoob MM, et al. Erythropoietin attenuates the tissue injury associated with hemorrhagic shock and myocardial ischemia. Shock. 2004;22(1):63–69. - PubMed
-
- Abrous DN, Koehl M, Le Moal M. Adult neurogenesis: from precursors to network and physiology. Physiol Rev. 2005;85(2):523–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
