

Osteopontin is induced by hedgehog pathway activation and promotes fibrosis progression in nonalcoholic steatohepatitis
- PMID: 20967826
- PMCID: PMC3025083
- DOI: 10.1002/hep.23998
Osteopontin is induced by hedgehog pathway activation and promotes fibrosis progression in nonalcoholic steatohepatitis
Abstract
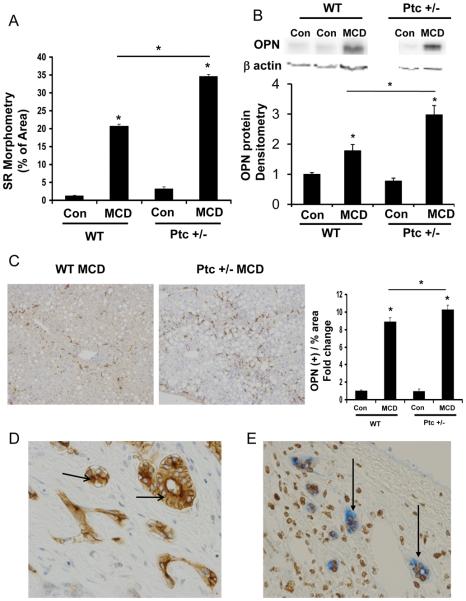
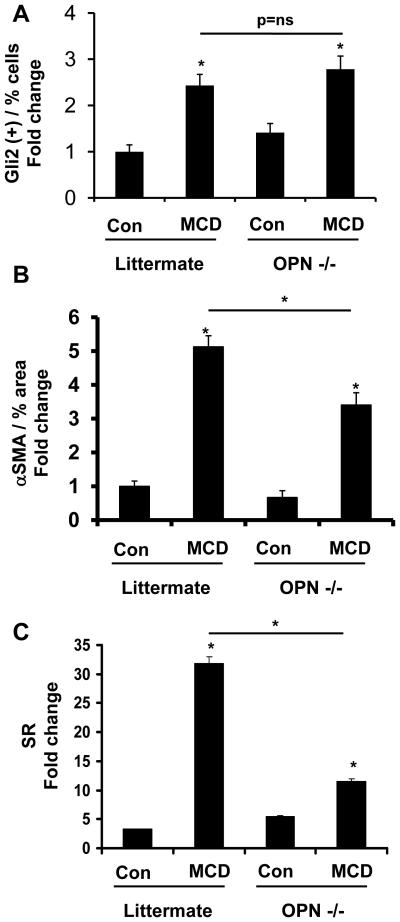
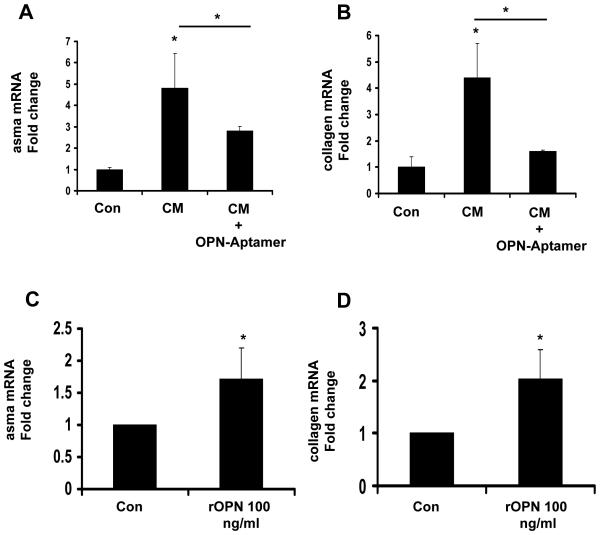
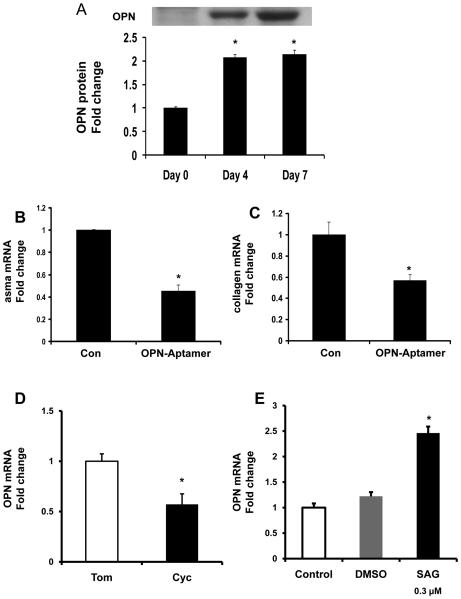
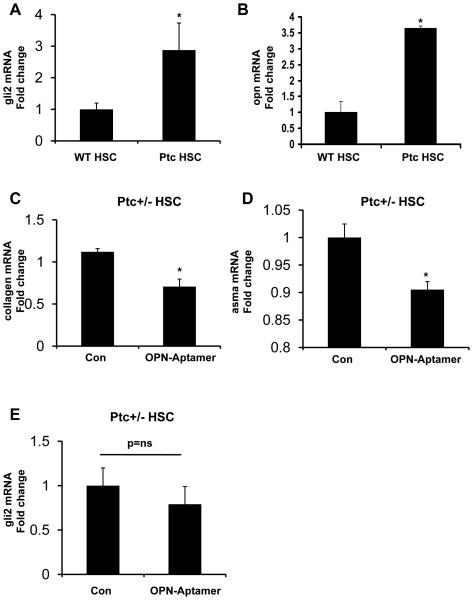
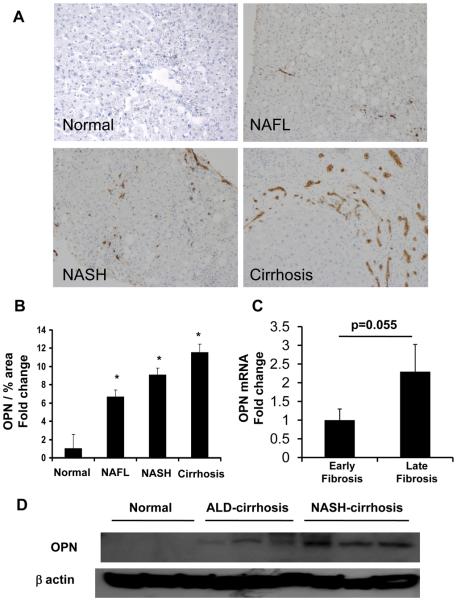
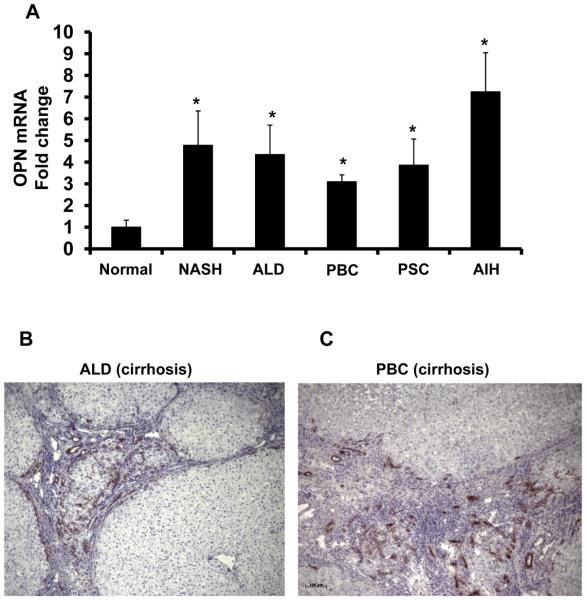
Nonalcoholic steatohepatitis (NASH) is a leading cause of cirrhosis. Recently, we showed that NASH-related cirrhosis is associated with Hedgehog (Hh) pathway activation. The gene encoding osteopontin (OPN), a profibrogenic extracellular matrix protein and cytokine, is a direct transcriptional target of the Hh pathway. Thus, we hypothesize that Hh signaling induces OPN to promote liver fibrosis in NASH. Hepatic OPN expression and liver fibrosis were analyzed in wild-type (WT) mice, Patched-deficient (Ptc(+/-) ) (overly active Hh signaling) mice, and OPN-deficient mice before and after feeding methionine and choline-deficient (MCD) diets to induce NASH-related fibrosis. Hepatic OPN was also quantified in human NASH and nondiseased livers. Hh signaling was manipulated in cultured liver cells to assess direct effects on OPN expression, and hepatic stellate cells (HSCs) were cultured in medium with different OPN activities to determine effects on HSC phenotype. When fed MCD diets, Ptc(+/-) mice expressed more OPN and developed worse liver fibrosis (P < 0.05) than WT mice, whereas OPN-deficient mice exhibited reduced fibrosis (P < 0.05). In NASH patients, OPN was significantly up-regulated and correlated with Hh pathway activity and fibrosis stage. During NASH, ductular cells strongly expressed OPN. In cultured HSCs, SAG (an Hh agonist) up-regulated, whereas cyclopamine (an Hh antagonist) repressed OPN expression (P < 0.005). Cholangiocyte-derived OPN and recombinant OPN promoted fibrogenic responses in HSCs (P < 0.05); neutralizing OPN with RNA aptamers attenuated this (P < 0.05).
Conclusion: OPN is Hh-regulated and directly promotes profibrogenic responses. OPN induction correlates with Hh pathway activity and fibrosis stage. Therefore, OPN inhibition may be beneficial in NASH.
Copyright © 2010 American Association for the Study of Liver Diseases.
Figures
Comment in
-
Osteopontin: a missing link between hedgehog signaling and fibrosis in nonalcoholic steatohepatitis.Hepatology. 2011 Feb;53(2):382-4. doi: 10.1002/hep.24118. Hepatology. 2011. PMID: 21274859 No abstract available.
References
-
- Feldstein AE, Canbay A, Angulo P, Taniai M, Burgart LJ, Lindor KD, Gores GJ. Hepatocyte apoptosis and fas expression are prominent features of human nonalcoholic steatohepatitis. Gastroenterology. 2003;125:437–443. - PubMed
-
- Wieckowska A, Zein NN, Yerian LM, Lopez AR, McCullough AJ, Feldstein AE. In vivo assessment of liver cell apoptosis as a novel biomarker of disease severity in nonalcoholic fatty liver disease. Hepatology. 2006;44:27–33. - PubMed
-
- Feldstein AE, Papouchado BG, Angulo P, Sanderson S, Adams L, Gores GJ. Hepatic stellate cells and fibrosis progression in patients with nonalcoholic fatty liver disease. Clin Gastroenterol Hepatol. 2005;3:384–389. - PubMed
-
- Choi SS, Omenetti A, Witek RP, Moylan CA, Syn WK, Jung Y, Yang L, et al. Hedgehog pathway activation and epithelial-to-mesenchymal transitions during myofibroblastic transformation of rat hepatic cells in culture and cirrhosis. Am J Physiol Gastrointest Liver Physiol. 2009;297:G1093–1106. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials