

Rec8-containing cohesin maintains bivalents without turnover during the growing phase of mouse oocytes
- PMID: 20971813
- PMCID: PMC2975927
- DOI: 10.1101/gad.605910
Rec8-containing cohesin maintains bivalents without turnover during the growing phase of mouse oocytes
Abstract
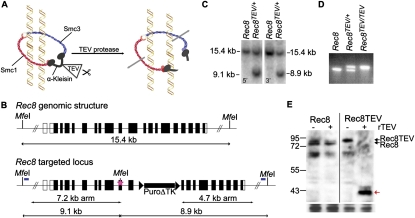
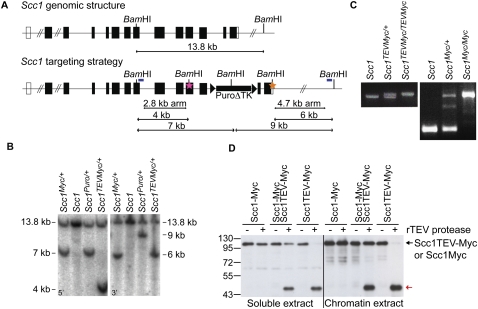
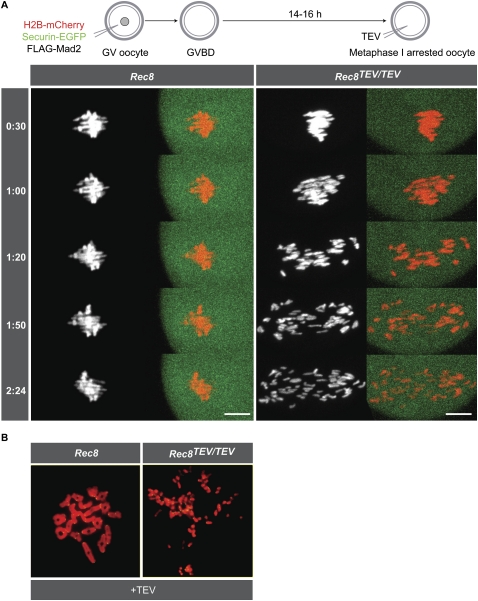
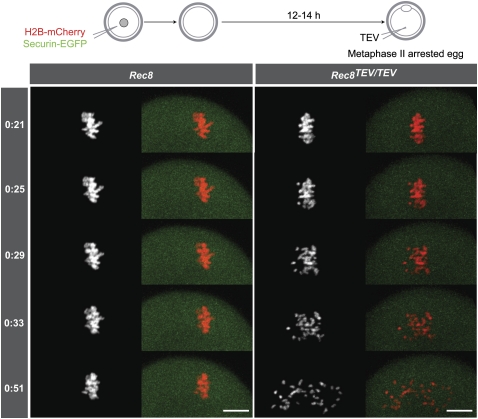
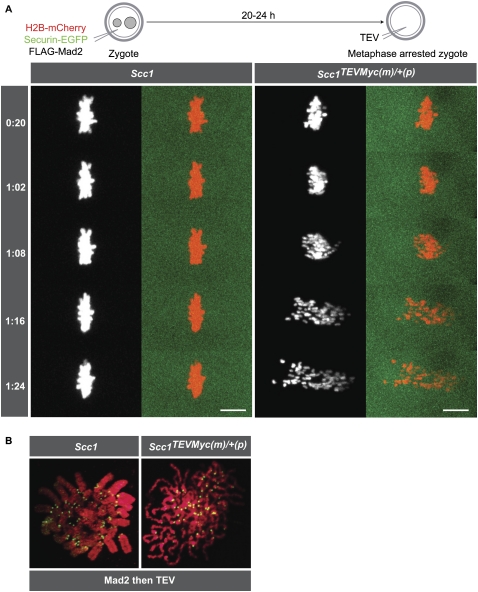
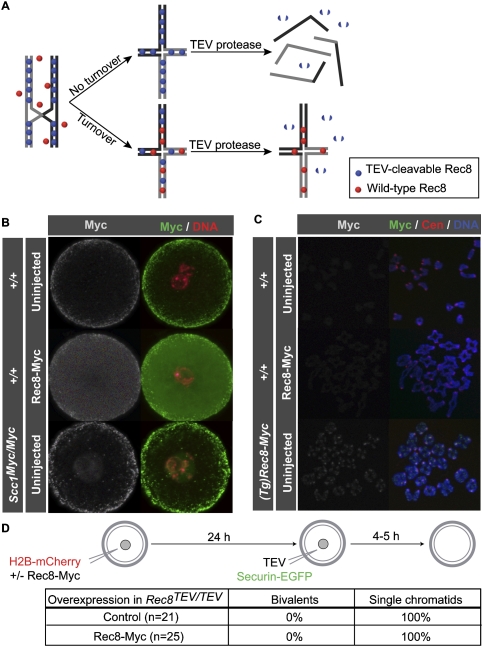
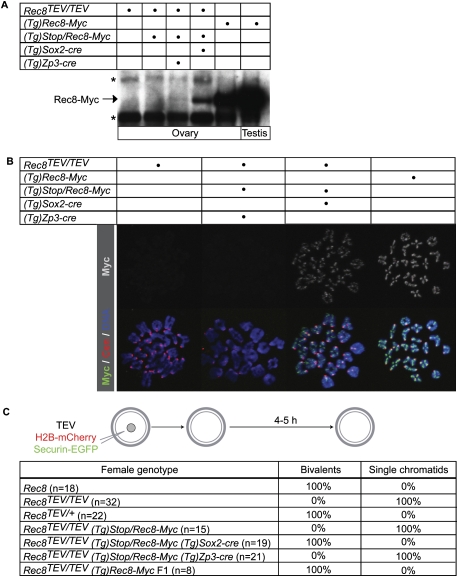
During female meiosis, bivalent chromosomes are thought to be held together from birth until ovulation by sister chromatid cohesion mediated by cohesin complexes whose ring structure depends on kleisin subunits, either Rec8 or Scc1. Because cohesion is established at DNA replication in the embryo, its maintenance for such a long time may require cohesin turnover. To address whether Rec8- or Scc1-containing cohesin holds bivalents together and whether it turns over, we created mice whose kleisin subunits can be cleaved by TEV protease. We show by microinjection experiments and confocal live-cell imaging that Rec8 cleavage triggers chiasmata resolution during meiosis I and sister centromere disjunction during meiosis II, while Scc1 cleavage triggers sister chromatid disjunction in the first embryonic mitosis, demonstrating a dramatic transition from Rec8- to Scc1-containing cohesin at fertilization. Crucially, activation of an ectopic Rec8 transgene during the growing phase of Rec8(TEV)(/TEV) oocytes does not prevent TEV-mediated bivalent destruction, implying little or no cohesin turnover for ≥2 wk during oocyte growth. We suggest that the inability of oocytes to regenerate cohesion may contribute to age-related meiosis I errors.
Figures
Comment in
-
Deterioration without replenishment--the misery of oocyte cohesin.Genes Dev. 2010 Dec 1;24(23):2587-91. doi: 10.1101/gad.2000610. Genes Dev. 2010. PMID: 21123645 Free PMC article.
References
-
- Adams DJ, Quail MA, Cox T, van der Weyden L, Gorick BD, Su Q, Chan WI, Davies R, Bonfield JK, Law F, et al. 2005. A genome-wide, end-sequenced 129Sv BAC library resource for targeting vector construction. Genomics 86: 753–758 - PubMed
-
- Bannister LA, Reinhold LG, Munroe RJ, Schimenti JC 2004. Positional cloning and characterization of mouse mei8, a disrupted allele of the meiotic cohesin Rec8. Genesis 40: 184–194 - PubMed
-
- Buonomo SB, Clyne RK, Fuchs J, Loidl J, Uhlmann F, Nasmyth K 2000. Disjunction of homologous chromosomes in meiosis I depends on proteolytic cleavage of the meiotic cohesin Rec8 by separin. Cell 103: 387–398 - PubMed
-
- Días-Martínez LA, Giménez-Abián JF, Clarke DJ 2008. Chromosome cohesion—Rings, knots, orcs and fellowship. J Cell Sci 121: 2107–2114 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases