

Plantazolicin, a novel microcin B17/streptolysin S-like natural product from Bacillus amyloliquefaciens FZB42
- PMID: 20971906
- PMCID: PMC3019963
- DOI: 10.1128/JB.00784-10
Plantazolicin, a novel microcin B17/streptolysin S-like natural product from Bacillus amyloliquefaciens FZB42
Abstract
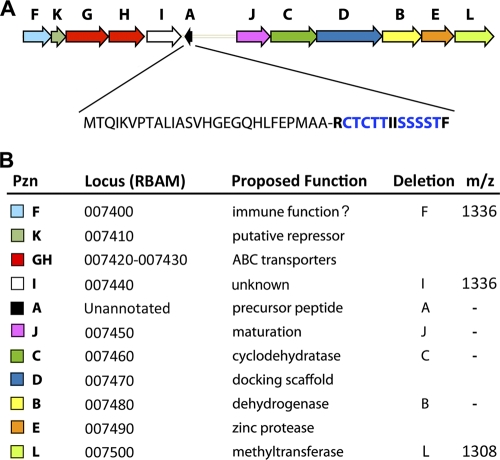
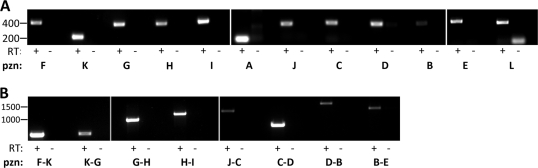
Here we report on a novel thiazole/oxazole-modified microcin (TOMM) from Bacillus amyloliquefaciens FZB42, a Gram-positive soil bacterium. This organism is well known for stimulating plant growth and biosynthesizing complex small molecules that suppress the growth of bacterial and fungal plant pathogens. Like microcin B17 and streptolysin S, the TOMM from B. amyloliquefaciens FZB42 undergoes extensive posttranslational modification to become a bioactive natural product. Our data show that the modified peptide bears a molecular mass of 1,335 Da and displays antibacterial activity toward closely related Gram-positive bacteria. A cluster of 12 genes that covers ∼10 kb is essential for the production, modification, export, and self-immunity of this natural product. We have named this compound plantazolicin (PZN), based on the association of several producing organisms with plants and the incorporation of azole heterocycles, which derive from Cys, Ser, and Thr residues of the precursor peptide.
Figures
Similar articles
-
Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42.J Bacteriol. 2014 May;196(10):1842-52. doi: 10.1128/JB.01474-14. Epub 2014 Mar 7. J Bacteriol. 2014. PMID: 24610713 Free PMC article.
-
The highly modified microcin peptide plantazolicin is associated with nematicidal activity of Bacillus amyloliquefaciens FZB42.Appl Microbiol Biotechnol. 2013 Dec;97(23):10081-90. doi: 10.1007/s00253-013-5247-5. Epub 2013 Oct 2. Appl Microbiol Biotechnol. 2013. PMID: 24085393
-
Structure determination and interception of biosynthetic intermediates for the plantazolicin class of highly discriminating antibiotics.ACS Chem Biol. 2011 Dec 16;6(12):1307-13. doi: 10.1021/cb200339d. Epub 2011 Oct 6. ACS Chem Biol. 2011. PMID: 21950656 Free PMC article.
-
The Microbial Toxin Microcin B17: Prospects for the Development of New Antibacterial Agents.J Mol Biol. 2019 Aug 23;431(18):3400-3426. doi: 10.1016/j.jmb.2019.05.050. Epub 2019 Jun 8. J Mol Biol. 2019. PMID: 31181289 Free PMC article. Review.
-
[Microcins--peptide antibiotics of enterobacteria: genetic control of the synthesis, structure, and mechanism of action].Genetika. 1999 Jan;35(1):5-16. Genetika. 1999. PMID: 10330606 Review. Russian.
Cited by
-
Biocontrol properties from phyllospheric bacteria isolated from Solanum lycopersicum and Lactuca sativa and genome mining of antimicrobial gene clusters.BMC Genomics. 2022 Feb 21;23(1):152. doi: 10.1186/s12864-022-08392-0. BMC Genomics. 2022. PMID: 35189837 Free PMC article.
-
Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42.J Bacteriol. 2014 May;196(10):1842-52. doi: 10.1128/JB.01474-14. Epub 2014 Mar 7. J Bacteriol. 2014. PMID: 24610713 Free PMC article.
-
The Characterization, Biological Activities, and Potential Applications of the Antimicrobial Peptides Derived from Bacillus spp.: A Comprehensive Review.Probiotics Antimicrob Proteins. 2025 Jun;17(3):1624-1647. doi: 10.1007/s12602-024-10447-5. Epub 2024 Dec 30. Probiotics Antimicrob Proteins. 2025. PMID: 39739161 Review.
-
Antimicrobial peptides from Bacillus spp. and strategies to enhance their yield.Appl Microbiol Biotechnol. 2023 Sep;107(18):5569-5593. doi: 10.1007/s00253-023-12651-9. Epub 2023 Jul 14. Appl Microbiol Biotechnol. 2023. PMID: 37450018 Review.
-
A Seed-Endophytic Bacillus safensis Strain With Antimicrobial Activity Has Genes for Novel Bacteriocin-Like Antimicrobial Peptides.Front Microbiol. 2021 Sep 27;12:734216. doi: 10.3389/fmicb.2021.734216. eCollection 2021. Front Microbiol. 2021. PMID: 34646254 Free PMC article.
References
-
- Borriss, R., X. Chen, C. Rueckert, J. Blom, A. Becker, B. Baumgarth, B. Fan, R. Pukall, P. Schumann, C. Sproer, H. Junge, J. Vater, A. Puhler, and H. P. Klenk. 3 September 2010. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM7T and FZB42: a proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on their discriminating complete genome sequences. Int. J. Syst. Evol. Microbiol. [Epub ahead of print.] doi:10.1099/ijs.0.023267-0. - DOI - PubMed
-
- Busenlehner, L. S., M. A. Pennella, and D. P. Giedroc. 2003. The SmtB/ArsR family of metalloregulatory transcriptional repressors: structural insights into prokaryotic metal resistance. FEMS Microbiol. Rev. 27:131-143. - PubMed
-
- Butcher, B. G., and J. D. Helmann. 2006. Identification of Bacillus subtilis sigma-dependent genes that provide intrinsic resistance to antimicrobial compounds produced by Bacilli. Mol. Microbiol. 60:765-782. - PubMed
-
- Chen, X. 2009. Whole genome analysis of the plant growth-promoting rhizobacteria Bacillus amyloliquefaciens FZB42 with focus on its secondary metabolites. Ph.D. dissertation. Humboldt-Universität zu Berlin, Berlin, Germany.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
