

Conservation and divergence of autonomous pathway genes in the flowering regulatory network of Beta vulgaris
- PMID: 20974738
- PMCID: PMC3130164
- DOI: 10.1093/jxb/erq321
Conservation and divergence of autonomous pathway genes in the flowering regulatory network of Beta vulgaris
Abstract
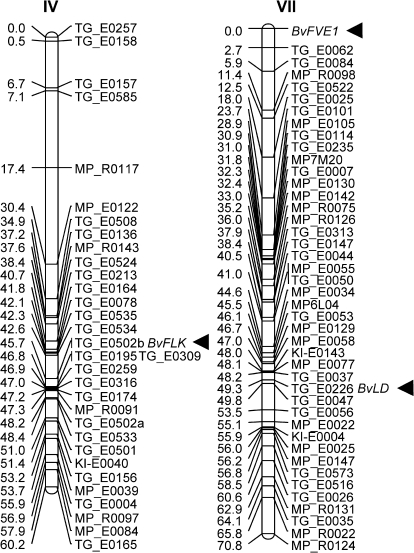
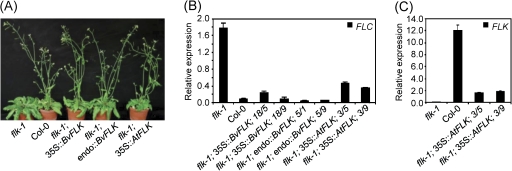
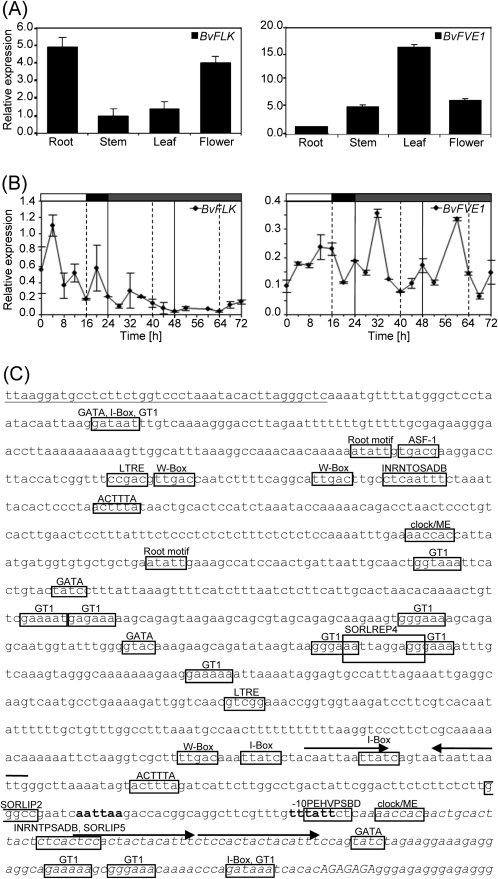
The transition from vegetative growth to reproductive development is a complex process that requires an integrated response to multiple environmental cues and endogenous signals. In Arabidopsis thaliana, which has a facultative requirement for vernalization and long days, the genes of the autonomous pathway function as floral promoters by repressing the central repressor and vernalization-regulatory gene FLC. Environmental regulation by seasonal changes in daylength is under control of the photoperiod pathway and its key gene CO. The root and leaf crop species Beta vulgaris in the caryophyllid clade of core eudicots, which is only very distantly related to Arabidopsis, is an obligate long-day plant and includes forms with or without vernalization requirement. FLC and CO homologues with related functions in beet have been identified, but the presence of autonomous pathway genes which function in parallel to the vernalization and photoperiod pathways has not yet been reported. Here, this begins to be addressed by the identification and genetic mapping of full-length homologues of the RNA-regulatory gene FLK and the chromatin-regulatory genes FVE, LD, and LDL1. When overexpressed in A. thaliana, BvFLK accelerates bolting in the Col-0 background and fully complements the late-bolting phenotype of an flk mutant through repression of FLC. In contrast, complementation analysis of BvFVE1 and the presence of a putative paralogue in beet suggest evolutionary divergence of FVE homologues. It is further shown that BvFVE1, unlike FVE in Arabidopsis, is under circadian clock control. Together, the data provide first evidence for evolutionary conservation of components of the autonomous pathway in B. vulgaris, while also suggesting divergence or subfunctionalization of one gene. The results are likely to be of broader relevance because B. vulgaris expands the spectrum of evolutionarily diverse species which are subject to differential developmental and/or environmental regulation of floral transition.
Figures

References
-
- Abegg FA. A genetic factor for the annual habit in beets and linkage relationship. Journal of Agricultural Research. 1936;53:493–511.
-
- Alonso JM, Stepanova AN, Leisse TJ, et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003;301:653–657. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers E, Lipman DJ. Basic local alignment search tool. Journal of Molecular Biology. 1990;215:403–410. - PubMed
-
- Amasino R. Take a cold flower. Nature Genetics. 2004;36:111–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
