

The stearoyl-coenzyme A desaturase 1 is essential for virulence and membrane stress in Candida parapsilosis through unsaturated fatty acid production
- PMID: 20974817
- PMCID: PMC3019882
- DOI: 10.1128/IAI.00753-10
The stearoyl-coenzyme A desaturase 1 is essential for virulence and membrane stress in Candida parapsilosis through unsaturated fatty acid production
Abstract
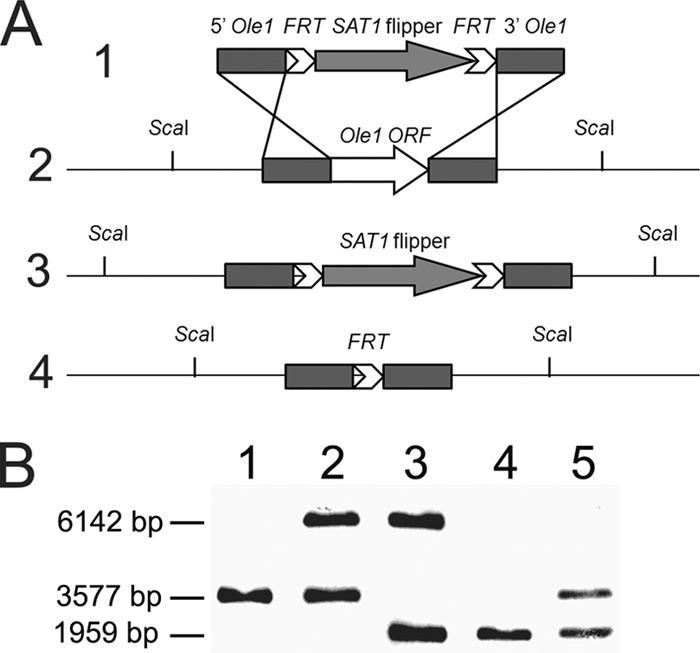
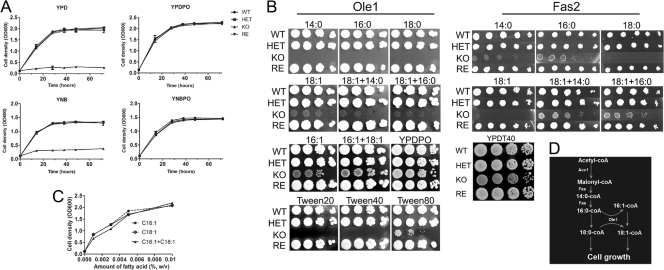
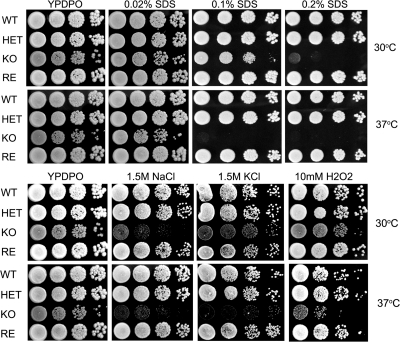
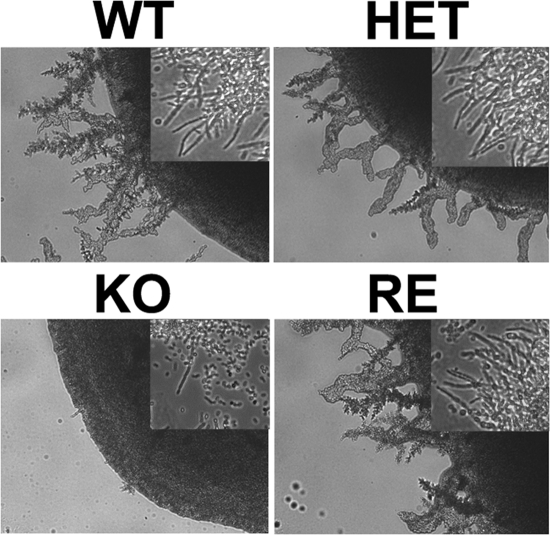
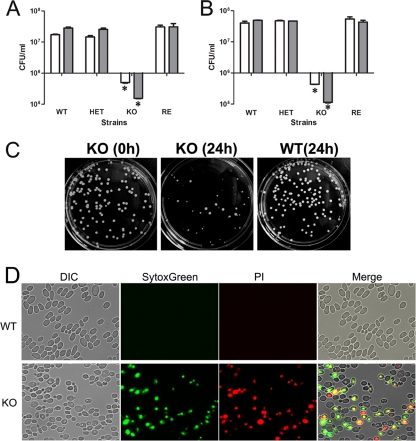
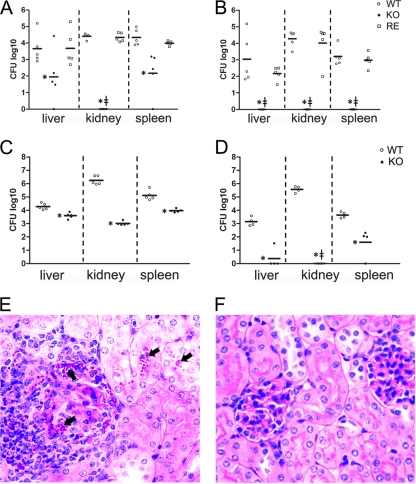
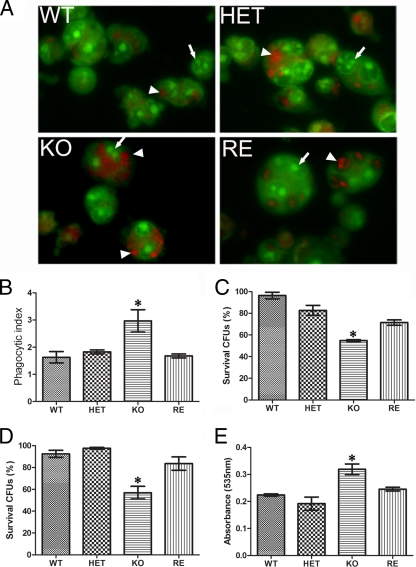
Unsaturated fatty acids (UFA) are essential components of cells. In Saccharomyces cerevisiae, stearoyl-coenzyme A (CoA) desaturase 1 (OLE1) affects cell viability through the regulation of oleic (18:1) or palmitoleic (16:1) acid production. In this study, we used a targeted gene deletion approach to determine the impact of OLE1 on the emerging human pathogenic fungus Candida parapsilosis. We found that the deletion of OLE1 resulted in an auxotrophic yeast strain (designated OLE1 KO) that required unsaturated fatty acids for growth but not saturated fatty acids. Additionally, the production of UFA by OLE1 KO yeast cells was markedly reduced, suggesting that Ole1 is essential for UFA production. In contrast to wild-type C. parapsilosis, which produced pseudohyphal growth on UFA-supplemented medium agar, pseudohyphal formation in the OLE1 KO cells was severely impaired, suggesting that Ole1 regulates morphology. Furthermore, the OLE1 KO cells were hypersensitive to various stress-inducing factors, such as salts, SDS, and H(2)O(2), especially at the physiological temperature. The results indicate that OLE1 is essential for the stress response, perhaps through the production of UFA for cell membrane biosynthesis. The OLE1 KO cells also were hypersensitive to human and fetal bovine serum, suggesting that targeting Ole1 could suppress the dissemination of yeast cells in the bloodstream. Murine-like macrophage J774.16 more efficiently killed the OLE1 KO yeasts, and significantly larger amounts of nitric oxide were detected in cocultures of macrophages and OLE1 KO cells than with wild-type or heterozygous strains. Moreover, the disruption of OLE1 significantly reduced fungal virulence in systemic murine infection. Taken together, these results demonstrate that Ole1 regulates the pathobiology of C. parapsilosis via UFA and that the OLE1 pathway is a promising antifungal target.
Figures
Similar articles
-
Chemical genetic profiling and characterization of small-molecule compounds that affect the biosynthesis of unsaturated fatty acids in Candida albicans.J Biol Chem. 2009 Jul 17;284(29):19754-64. doi: 10.1074/jbc.M109.019877. Epub 2009 Jun 1. J Biol Chem. 2009. PMID: 19487691 Free PMC article.
-
Regulatory elements that control transcription activation and unsaturated fatty acid-mediated repression of the Saccharomyces cerevisiae OLE1 gene.J Biol Chem. 1996 Feb 16;271(7):3581-9. doi: 10.1074/jbc.271.7.3581. J Biol Chem. 1996. PMID: 8631965
-
Molecular cloning and characterization of the Delta9 fatty acid desaturase gene and its promoter region from Saccharomyces kluyveri.FEMS Yeast Res. 2002 Aug;2(3):333-9. doi: 10.1016/S1567-1356(02)00088-0. FEMS Yeast Res. 2002. PMID: 12702283
-
Fungal Δ9-fatty acid desaturase: a unique enzyme at the core of lipid metabolism in Aspergillus fumigatus and a promising target for the search for antifungal strategies.mBio. 2025 Aug 13;16(8):e0080324. doi: 10.1128/mbio.00803-24. Epub 2025 Jun 26. mBio. 2025. PMID: 40569070 Free PMC article. Review.
-
Yeast desaturases.Biochem Soc Trans. 2002 Nov;30(Pt 6):1080-2. doi: 10.1042/bst0301080. Biochem Soc Trans. 2002. PMID: 12440977 Review.
Cited by
-
HapX, an Indispensable bZIP Transcription Factor for Iron Acquisition, Regulates Infection Initiation by Orchestrating Conidial Oleic Acid Homeostasis and Cytomembrane Functionality in Mycopathogen Beauveria bassiana.mSystems. 2020 Oct 13;5(5):e00695-20. doi: 10.1128/mSystems.00695-20. mSystems. 2020. PMID: 33051379 Free PMC article.
-
Membrane fluidity and temperature sensing are coupled via circuitry comprised of Ole1, Rsp5, and Hsf1 in Candida albicans.Eukaryot Cell. 2014 Aug;13(8):1077-84. doi: 10.1128/EC.00138-14. Epub 2014 Jun 20. Eukaryot Cell. 2014. PMID: 24951438 Free PMC article.
-
Three, two, one yeast fatty acid desaturases: regulation and function.World J Microbiol Biotechnol. 2017 May;33(5):89. doi: 10.1007/s11274-017-2257-y. Epub 2017 Apr 7. World J Microbiol Biotechnol. 2017. PMID: 28390014 Review.
-
Candida parapsilosis produces prostaglandins from exogenous arachidonic acid and OLE2 is not required for their synthesis.Virulence. 2015;6(1):85-92. doi: 10.4161/21505594.2014.988097. Virulence. 2015. PMID: 25654274 Free PMC article.
-
A potent plant-derived antifungal acetylenic acid mediates its activity by interfering with fatty acid homeostasis.Antimicrob Agents Chemother. 2012 Jun;56(6):2894-907. doi: 10.1128/AAC.05663-11. Epub 2012 Mar 19. Antimicrob Agents Chemother. 2012. PMID: 22430960 Free PMC article.
References
-
- Bandyopadhyay, S., R. Zhan, Y. Wang, S. K. Pai, S. Hirota, S. Hosobe, Y. Takano, K. Saito, E. Furuta, M. Iiizumi, S. Mohinta, M. Watabe, C. Chalfant, and K. Watabe. 2006. Mechanism of apoptosis induced by the inhibition of fatty acid synthase in breast cancer cells. Cancer Res. 66:5934-5940. - PubMed
-
- Brinster, S., G. Lamberet, B. Staels, P. Trieu-Cuot, A. Gruss, and C. Poyart. 2009. Type II fatty acid synthesis is not a suitable antibiotic target for Gram-positive pathogens. Nature 458:83-86. - PubMed
-
- Chellappa, R., P. Kandasamy, C. S. Oh, Y. Jiang, M. Vemula, and C. E. Martin. 2001. The membrane proteins, Spt23p and Mga2p, play distinct roles in the activation of Saccharomyces cerevisiae OLE1 gene expression. Fatty acid-mediated regulation of Mga2p activity is independent of its proteolytic processing into a soluble transcription activator. J. Biol. Chem. 276:43548-43556. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
