

C4GEM, a genome-scale metabolic model to study C4 plant metabolism
- PMID: 20974891
- PMCID: PMC2996019
- DOI: 10.1104/pp.110.166488
C4GEM, a genome-scale metabolic model to study C4 plant metabolism
Abstract
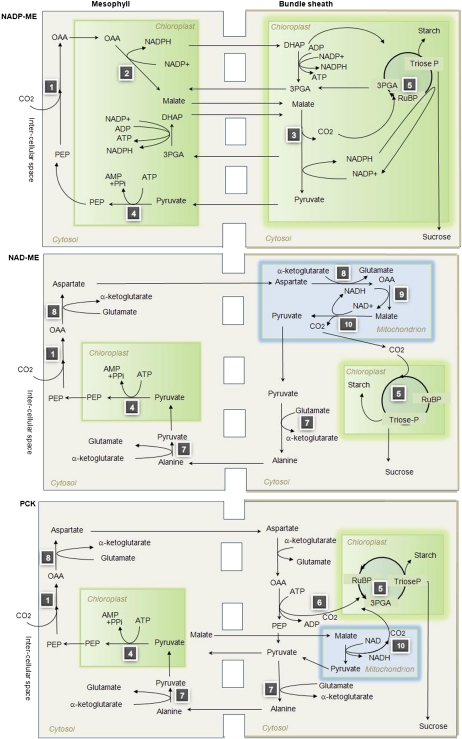
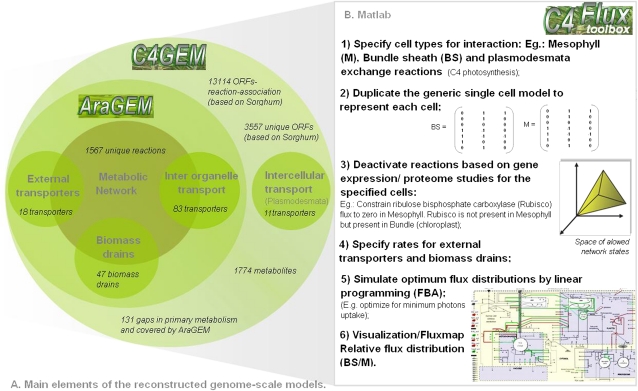
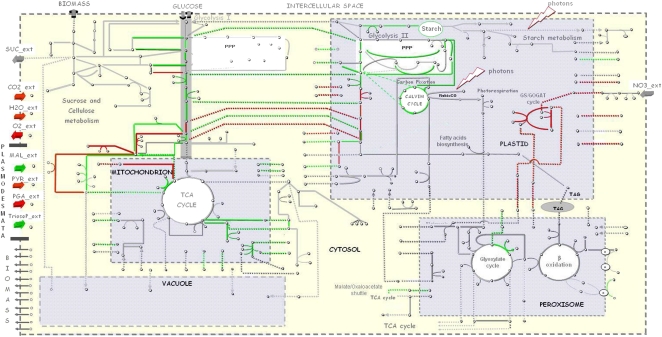
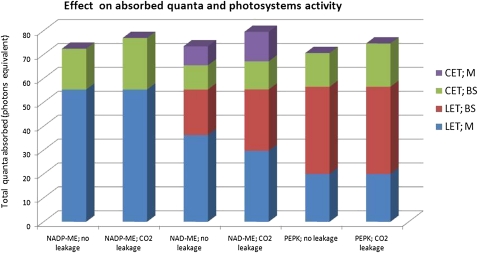
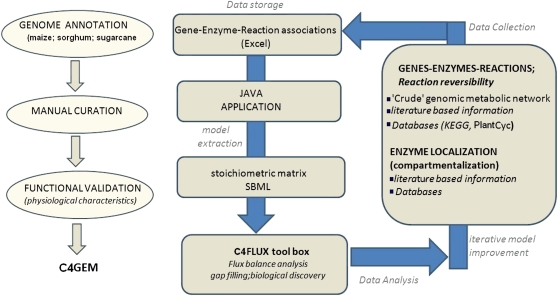
Leaves of C(4) grasses (such as maize [Zea mays], sugarcane [Saccharum officinarum], and sorghum [Sorghum bicolor]) form a classical Kranz leaf anatomy. Unlike C(3) plants, where photosynthetic CO(2) fixation proceeds in the mesophyll (M), the fixation process in C(4) plants is distributed between two cell types, the M cell and the bundle sheath (BS) cell. Here, we develop a C(4) genome-scale model (C4GEM) for the investigation of flux distribution in M and BS cells during C(4) photosynthesis. C4GEM, to our knowledge, is the first large-scale metabolic model that encapsulates metabolic interactions between two different cell types. C4GEM is based on the Arabidopsis (Arabidopsis thaliana) model (AraGEM) but has been extended by adding reactions and transporters responsible to represent three different C(4) subtypes (NADP-ME [for malic enzyme], NAD-ME, and phosphoenolpyruvate carboxykinase). C4GEM has been validated for its ability to synthesize 47 biomass components and consists of 1,588 unique reactions, 1,755 metabolites, 83 interorganelle transporters, and 29 external transporters (including transport through plasmodesmata). Reactions in the common C(4) model have been associated with well-annotated C(4) species (NADP-ME subtypes): 3,557 genes in sorghum, 11,623 genes in maize, and 3,881 genes in sugarcane. The number of essential reactions not assigned to genes is 131, 135, and 156 in sorghum, maize, and sugarcane, respectively. Flux balance analysis was used to assess the metabolic activity in M and BS cells during C(4) photosynthesis. Our simulations were consistent with chloroplast proteomic studies, and C4GEM predicted the classical C(4) photosynthesis pathway and its major effect in organelle function in M and BS. The model also highlights differences in metabolic activities around photosystem I and photosystem II for three different C(4) subtypes. Effects of CO(2) leakage were also explored. C4GEM is a viable framework for in silico analysis of cell cooperation between M and BS cells during photosynthesis and can be used to explore C(4) plant metabolism.
Figures
References
-
- Agarie S, Miura A, Sumikura R, Tsukamoto S, Nose A, Arima S, Matsuoka M, Miyao-Tokutomi M. (2002) Overexpression of C4PEPC caused O2-insensitive photosynthesis in transgenic rice plants. Plant Sci 162: 257–265
-
- Bassi R, Marquardt J, Lavergne J. (1995) Biochemical and functional properties of photosystem II in agranal membranes from maize mesophyll and bundle sheath chloroplasts. Eur J Biochem 233: 709–719 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
