

Filament structure of bacterial tubulin homologue TubZ
- PMID: 20974911
- PMCID: PMC2993389
- DOI: 10.1073/pnas.1010176107
Filament structure of bacterial tubulin homologue TubZ
Abstract
Low copy number plasmids often depend on accurate partitioning systems for their continued survival. Generally, such systems consist of a centromere-like region of DNA, a DNA-binding adaptor, and a polymerizing cytomotive filament. Together these components drive newly replicated plasmids to opposite ends of the dividing cell. The Bacillus thuringiensis plasmid pBToxis relies on a filament of the tubulin/FtsZ-like protein TubZ for its segregation. By combining crystallography and electron microscopy, we have determined the structure of this filament. We explain how GTP hydrolysis weakens the subunit-subunit contact and also shed light on the partitioning of the plasmid-adaptor complex. The double helical superstructure of TubZ filaments is unusual for tubulin-like proteins. Filaments of ParM, the actin-like partitioning protein, are also double helical. We suggest that convergent evolution shapes these different types of cytomotive filaments toward a general mechanism for plasmid separation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
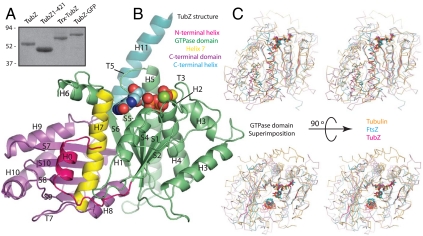
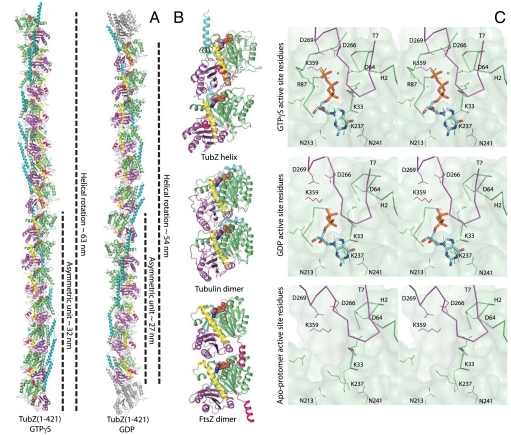

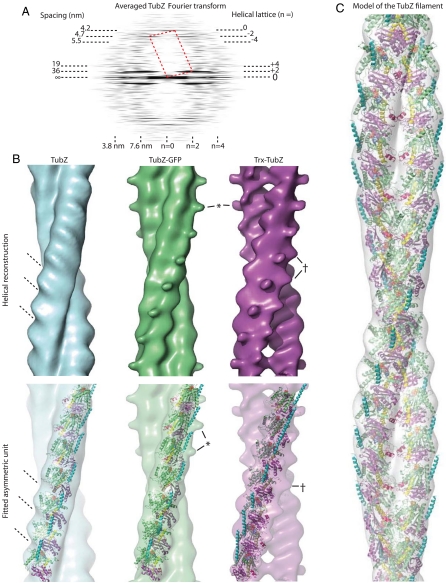
Comment in
-
When cytoskeletal worlds collide.Proc Natl Acad Sci U S A. 2010 Nov 16;107(46):19609-10. doi: 10.1073/pnas.1014665107. Epub 2010 Nov 8. Proc Natl Acad Sci U S A. 2010. PMID: 21059902 Free PMC article. No abstract available.
Similar articles
-
Reconstitution of a prokaryotic minus end-tracking system using TubRC centromeric complexes and tubulin-like protein TubZ filaments.Proc Natl Acad Sci U S A. 2015 Apr 14;112(15):E1845-50. doi: 10.1073/pnas.1423746112. Epub 2015 Mar 30. Proc Natl Acad Sci U S A. 2015. PMID: 25825718 Free PMC article.
-
Filament formation of the FtsZ/tubulin-like protein TubZ from the Bacillus cereus pXO1 plasmid.J Biol Chem. 2012 Sep 14;287(38):32103-12. doi: 10.1074/jbc.M112.373803. Epub 2012 Jul 30. J Biol Chem. 2012. PMID: 22847006 Free PMC article.
-
Bacterial tubulin TubZ-Bt transitions between a two-stranded intermediate and a four-stranded filament upon GTP hydrolysis.Proc Natl Acad Sci U S A. 2014 Mar 4;111(9):3407-12. doi: 10.1073/pnas.1318339111. Epub 2014 Feb 18. Proc Natl Acad Sci U S A. 2014. PMID: 24550513 Free PMC article.
-
Evolution of cytomotive filaments: the cytoskeleton from prokaryotes to eukaryotes.Int J Biochem Cell Biol. 2009 Feb;41(2):323-9. doi: 10.1016/j.biocel.2008.08.010. Epub 2008 Aug 13. Int J Biochem Cell Biol. 2009. PMID: 18768164 Review.
-
Tubulin-Like Proteins in Prokaryotic DNA Positioning.Subcell Biochem. 2017;84:323-356. doi: 10.1007/978-3-319-53047-5_11. Subcell Biochem. 2017. PMID: 28500531 Review.
Cited by
-
A novel transcriptional activator, tubX, is required for the stability of Bacillus sphaericus mosquitocidal plasmid pBsph.J Bacteriol. 2014 Dec;196(24):4304-14. doi: 10.1128/JB.01855-14. Epub 2014 Sep 29. J Bacteriol. 2014. PMID: 25266379 Free PMC article.
-
Plasmid Localization and Partition in Enterobacteriaceae.EcoSal Plus. 2019 Jun;8(2):10.1128/ecosalplus.ESP-0003-2019. doi: 10.1128/ecosalplus.ESP-0003-2019. EcoSal Plus. 2019. PMID: 31187729 Free PMC article. Review.
-
Management of cytoskeleton architecture by molecular chaperones and immunophilins.Cell Signal. 2011 Dec;23(12):1907-20. doi: 10.1016/j.cellsig.2011.07.023. Epub 2011 Aug 12. Cell Signal. 2011. PMID: 21864675 Free PMC article. Review.
-
Structural mechanism of ATP-induced polymerization of the partition factor ParF: implications for DNA segregation.J Biol Chem. 2012 Jul 27;287(31):26146-54. doi: 10.1074/jbc.M112.373696. Epub 2012 Jun 6. J Biol Chem. 2012. PMID: 22674577 Free PMC article.
-
Reconstitution of a prokaryotic minus end-tracking system using TubRC centromeric complexes and tubulin-like protein TubZ filaments.Proc Natl Acad Sci U S A. 2015 Apr 14;112(15):E1845-50. doi: 10.1073/pnas.1423746112. Epub 2015 Mar 30. Proc Natl Acad Sci U S A. 2015. PMID: 25825718 Free PMC article.
References
-
- Pogliano J. DNA segregation by bacterial actin homologs. Dev Cell. 2004;6:3–4. - PubMed
-
- Salje J. Plasmid segregation: How to survive as an extra piece of DNA. Crit Rev Biochem Mol Biol. 2010;45:296–317. - PubMed
-
- Gerdes K, Møller-Jensen J, Bugge Jensen R. Plasmid and chromosome partitioning: Surprises from phylogeny. Mol Microbiol. 2000;37:455–466. - PubMed
-
- Löwe J, Amos LA. Evolution of cytomotive filaments: The cytoskeleton from prokaryotes to eukaryotes. Int J Biochem Cell Biol. 2009;41:323–329. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
