

Plasticity and specificity of the circadian epigenome
- PMID: 20975756
- PMCID: PMC4071955
- DOI: 10.1038/nn.2668
Plasticity and specificity of the circadian epigenome
Abstract
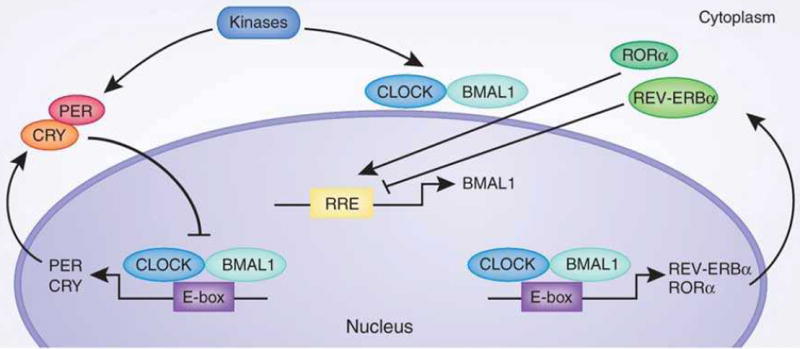
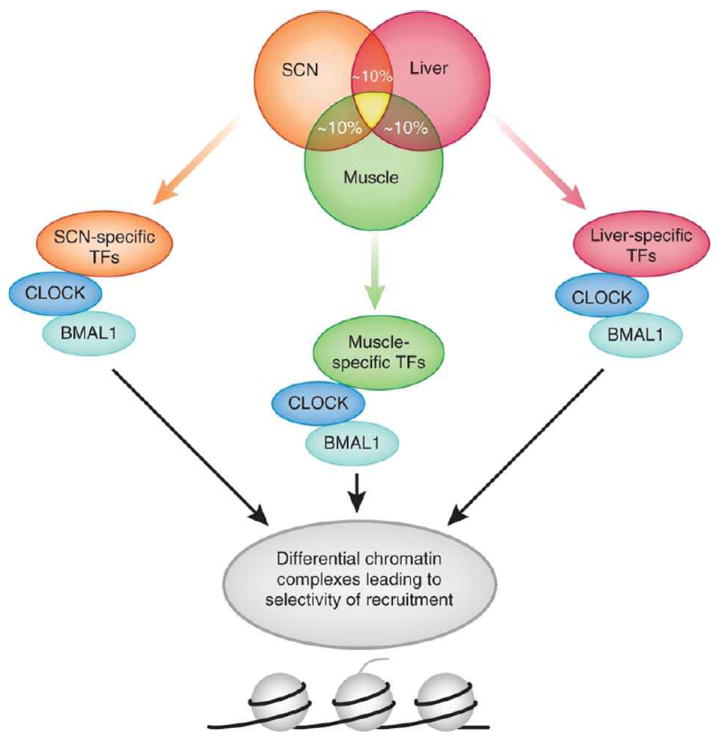
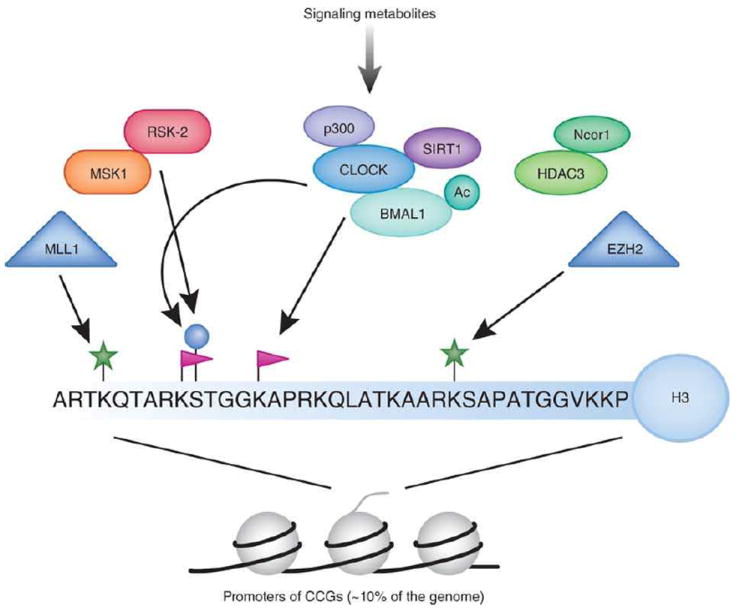
Circadian clocks control a variety of neuronal, behavioral and physiological responses, via transcriptional regulation of an appreciable portion of the genome. We describe the complex communication network between the brain-specific central clock and the tissue-specific peripheral clocks that serve to synchronize the organism to both external and internal demands. In addition, we discuss and speculate on how epigenetic processes are involved in creating transcriptional environments that are permissive to tissue-specific gene expression programs, which work in concert with the circadian machinery. Accumulating data show that chromatin remodeling events may be critical for providing specificity and plasticity in circadian regulation, and metabolic cues may be involved in directing such epigenetic events. A detailed understanding of the communication cues between the central and peripheral clocks is crucial for a more complete understanding of the circadian system and the several levels of control that are implicated in maintaining biological timekeeping.
Figures
References
-
- Reppert SM, Weaver DR. Molecular analysis of mammalian circadian rhythms. Annu Rev Physiol. 2001;63:647–76. - PubMed
-
- Bechtold DA, Gibbs JE, Loudon AS. Circadian dysfunction in disease. Trends Pharmacol Sci. 2010;31:191–8. - PubMed
-
- Sahar S, Sassone-Corsi P. Metabolism and cancer: the circadian clock connection. Nat Rev Cancer. 2009;9:886–96. - PubMed
-
- Thresher RJ, et al. Role of mouse cryptochrome blue-light photoreceptor in circadian photoresponses. Science. 1998;282:1490–4. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
