

Dual functions of ASCIZ in the DNA base damage response and pulmonary organogenesis
- PMID: 20975950
- PMCID: PMC2958817
- DOI: 10.1371/journal.pgen.1001170
Dual functions of ASCIZ in the DNA base damage response and pulmonary organogenesis
Erratum in
- PLoS Genet. 2010;6(11). doi: 10.1371/annotation/bce92e15-60b4-493b-89de-28529386f4ac doi: 10.1371/annotation/bce92e15-60b4-493b-89de-28529386f4ac
Abstract
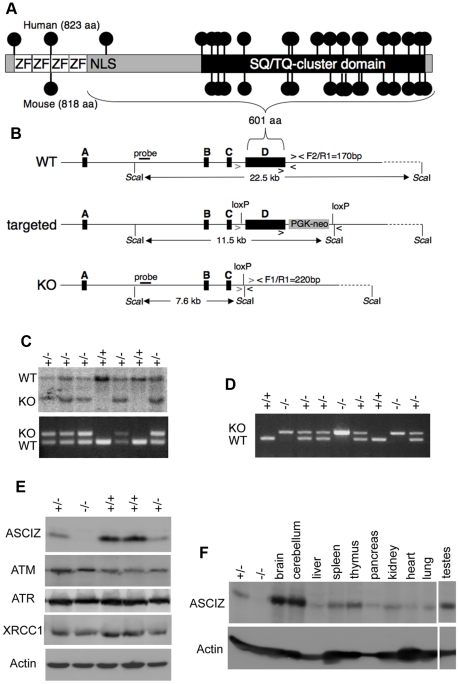
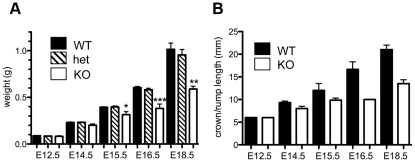
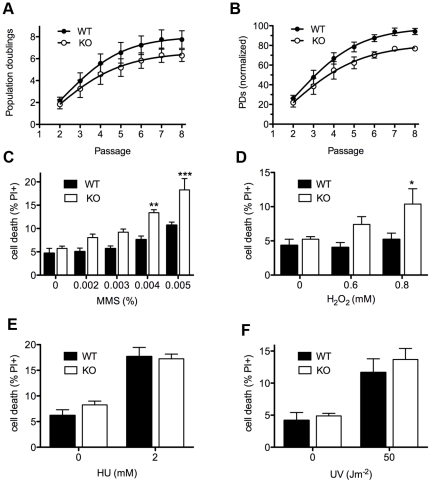
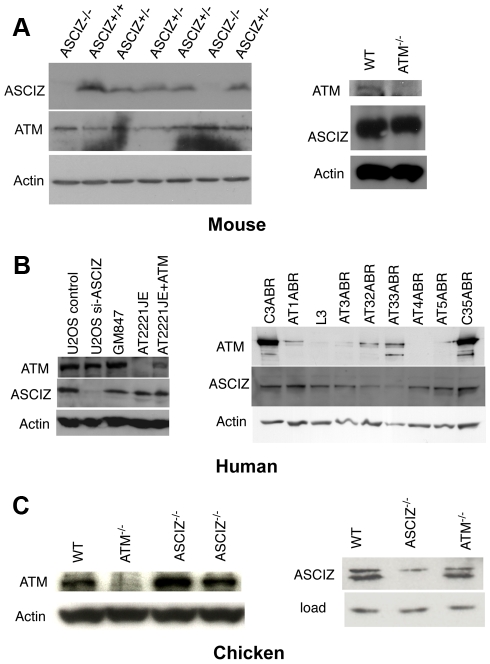
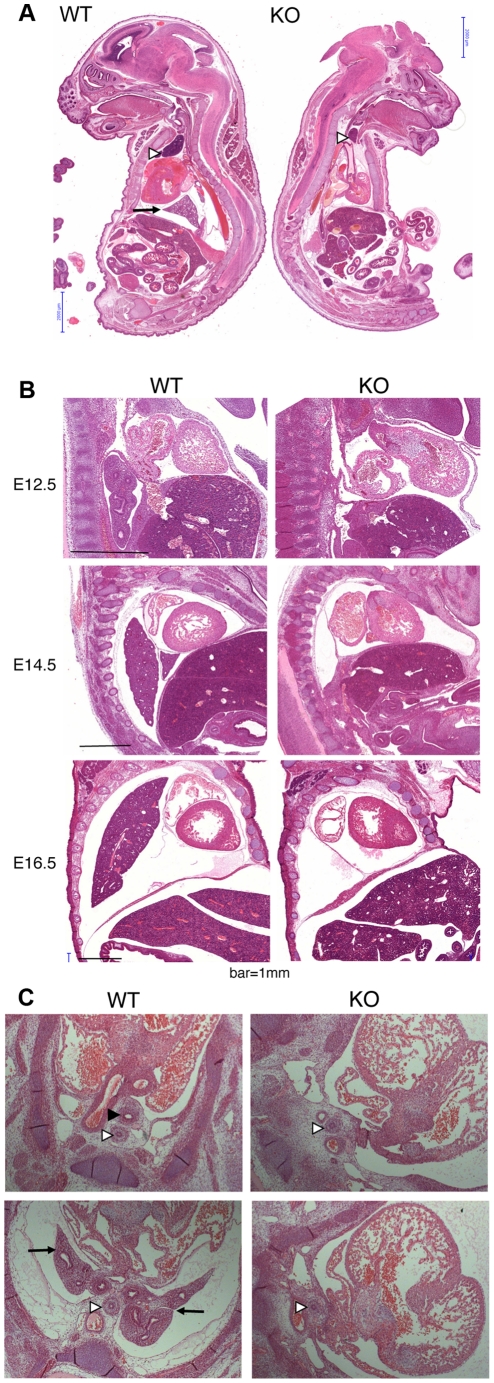
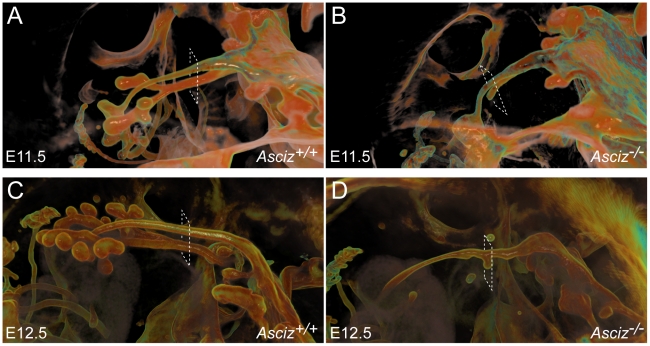
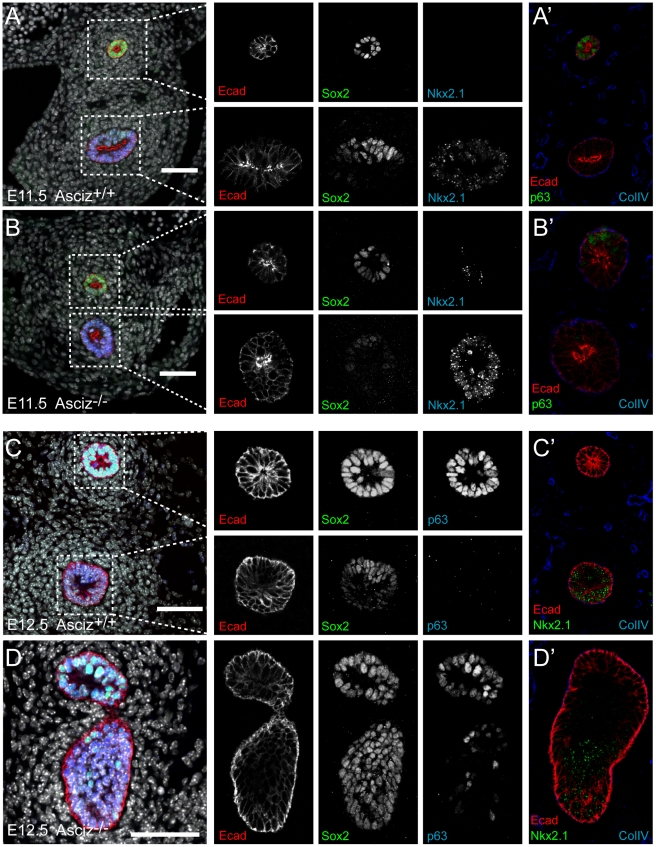
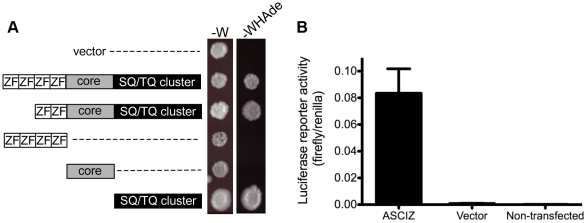
Zn²(+)-finger proteins comprise one of the largest protein superfamilies with diverse biological functions. The ATM substrate Chk2-interacting Zn²(+)-finger protein (ASCIZ; also known as ATMIN and ZNF822) was originally linked to functions in the DNA base damage response and has also been proposed to be an essential cofactor of the ATM kinase. Here we show that absence of ASCIZ leads to p53-independent late-embryonic lethality in mice. Asciz-deficient primary fibroblasts exhibit increased sensitivity to DNA base damaging agents MMS and H2O2, but Asciz deletion knock-down does not affect ATM levels and activation in mouse, chicken, or human cells. Unexpectedly, Asciz-deficient embryos also exhibit severe respiratory tract defects with complete pulmonary agenesis and severe tracheal atresia. Nkx2.1-expressing respiratory precursors are still specified in the absence of ASCIZ, but fail to segregate properly within the ventral foregut, and as a consequence lung buds never form and separation of the trachea from the oesophagus stalls early. Comparison of phenotypes suggests that ASCIZ functions between Wnt2-2b/ß-catenin and FGF10/FGF-receptor 2b signaling pathways in the mesodermal/endodermal crosstalk regulating early respiratory development. We also find that ASCIZ can activate expression of reporter genes via its SQ/TQ-cluster domain in vitro, suggesting that it may exert its developmental functions as a transcription factor. Altogether, the data indicate that, in addition to its role in the DNA base damage response, ASCIZ has separate developmental functions as an essential regulator of respiratory organogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
