

Telomere attrition occurs during ex vivo expansion of human dental pulp stem cells
- PMID: 20976265
- PMCID: PMC2952925
- DOI: 10.1155/2010/673513
Telomere attrition occurs during ex vivo expansion of human dental pulp stem cells
Abstract
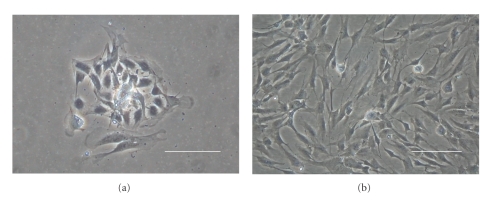
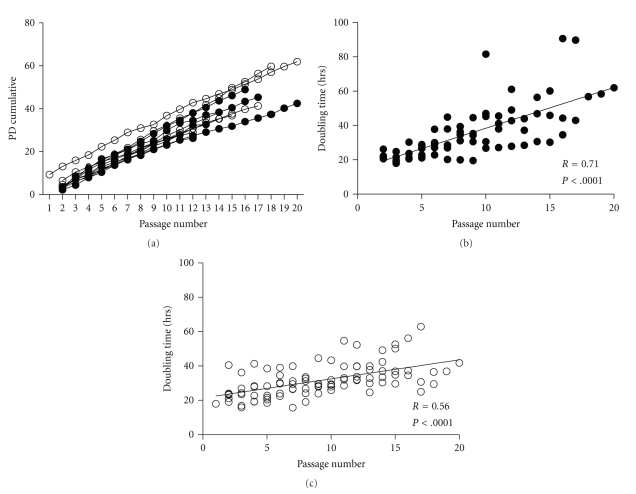
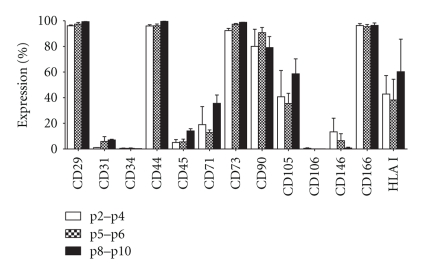
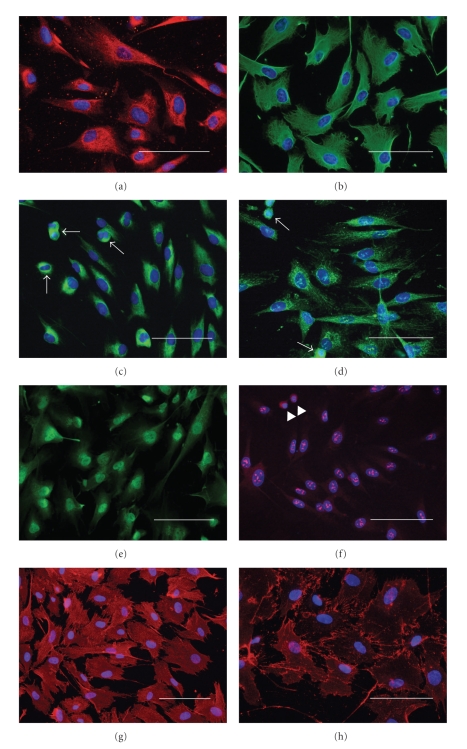
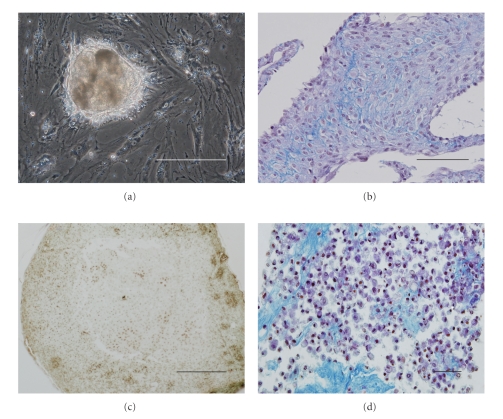
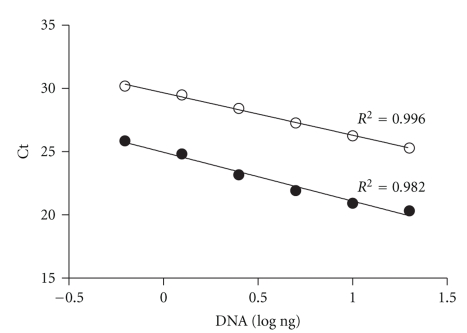
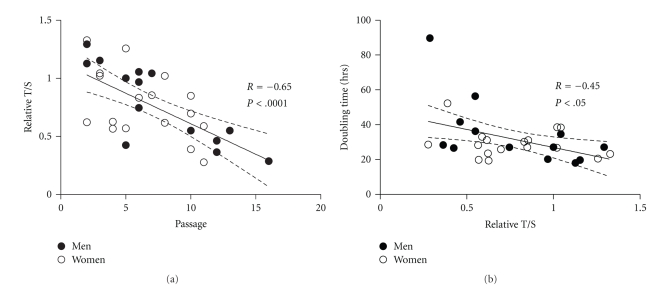
We provide a detailed characteristic of stem cells isolated and expanded from the human dental pulp. Dental pulp stem cells express mesenchymal cell markers STRO-1, vimentin, CD29, CD44, CD73, CD90, CD166, and stem cell markers Sox2, nestin, and nucleostemin. They are multipotent as shown by their osteogenic and chondrogenic potential. We measured relative telomere length in 11 dental pulp stem cell lines at different passages by quantitative real-time PCR. Despite their large proliferative capacity, stable viability, phenotype, and genotype over prolonged cultivation, human dental pulp stem cells suffer from progressive telomere shortening over time they replicate in vitro. Relative telomere length (T/S) was inversely correlated with cumulative doubling time. Our findings indicate that excessive ex vivo expansion of adult stem cells should be reduced at minimum to avoid detrimental effects on telomere maintenance and measurement of telomere length should become a standard when certificating the status and replicative age of stem cells prior therapeutic applications.
Figures
Similar articles
-
The Effect of Cultivation Passaging on the Relative Telomere Length and Proliferation Capacity of Dental Pulp Stem Cells.Biomolecules. 2021 Mar 20;11(3):464. doi: 10.3390/biom11030464. Biomolecules. 2021. PMID: 33804786 Free PMC article.
-
Mesenchymal stem cell characteristics of dental pulp and periodontal ligament stem cells after in vivo transplantation.Biomaterials. 2014 Aug;35(24):6332-43. doi: 10.1016/j.biomaterials.2014.04.071. Epub 2014 May 10. Biomaterials. 2014. PMID: 24824581
-
Isolation and prolonged expansion of oral mesenchymal stem cells under clinical-grade, GMP-compliant conditions differentially affects "stemness" properties.Stem Cell Res Ther. 2017 Nov 2;8(1):247. doi: 10.1186/s13287-017-0705-0. Stem Cell Res Ther. 2017. PMID: 29096714 Free PMC article.
-
Enzymatic Isolation, Amplification and Characterization of Dental Pulp Stem Cells.Folia Biol (Praha). 2019;65(3):124-133. doi: 10.14712/fb2019065030124. Folia Biol (Praha). 2019. PMID: 31638559
-
Carious deciduous teeth are a potential source for dental pulp stem cells.Clin Oral Investig. 2016 Jan;20(1):75-81. doi: 10.1007/s00784-015-1477-5. Epub 2015 Apr 22. Clin Oral Investig. 2016. PMID: 25898896
Cited by
-
Assessment of the Influence of Various Concentrations of Sodium Hypochlorite on Stem Cell Derived From Human Exfoliated Deciduous Teeth (SHED) Proliferation and Differentiation.Cureus. 2022 Dec 27;14(12):e33024. doi: 10.7759/cureus.33024. eCollection 2022 Dec. Cureus. 2022. PMID: 36726895 Free PMC article.
-
Usefulness of telomere length in DNA from human teeth for age estimation.Int J Legal Med. 2018 Mar;132(2):353-359. doi: 10.1007/s00414-017-1595-5. Epub 2017 Apr 24. Int J Legal Med. 2018. PMID: 28439695
-
Advances and Perspectives in Dental Pulp Stem Cell Based Neuroregeneration Therapies.Int J Mol Sci. 2021 Mar 29;22(7):3546. doi: 10.3390/ijms22073546. Int J Mol Sci. 2021. PMID: 33805573 Free PMC article. Review.
-
Tissue non-specific alkaline phosphatase production by human dental pulp stromal cells is enhanced by high density cell culture.Cell Tissue Res. 2015 Aug;361(2):529-40. doi: 10.1007/s00441-014-2106-3. Epub 2015 Feb 1. Cell Tissue Res. 2015. PMID: 25636587 Free PMC article.
-
The dual role of asporin in breast cancer progression.Oncotarget. 2016 Aug 9;7(32):52045-52060. doi: 10.18632/oncotarget.10471. Oncotarget. 2016. PMID: 27409832 Free PMC article.
References
-
- Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292(5819):154–156. - PubMed
-
- Strelchenko N, Verlinsky O, Kukharenko V, Verlinsky Y. Morula-derived human embryonic stem cells. Reproductive BioMedicine Online. 2004;9(6):623–629. - PubMed
-
- Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276(5309):71–74. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
