

Hox11 genes establish synovial joint organization and phylogenetic characteristics in developing mouse zeugopod skeletal elements
- PMID: 20978074
- PMCID: PMC3049277
- DOI: 10.1242/dev.053447
Hox11 genes establish synovial joint organization and phylogenetic characteristics in developing mouse zeugopod skeletal elements
Abstract
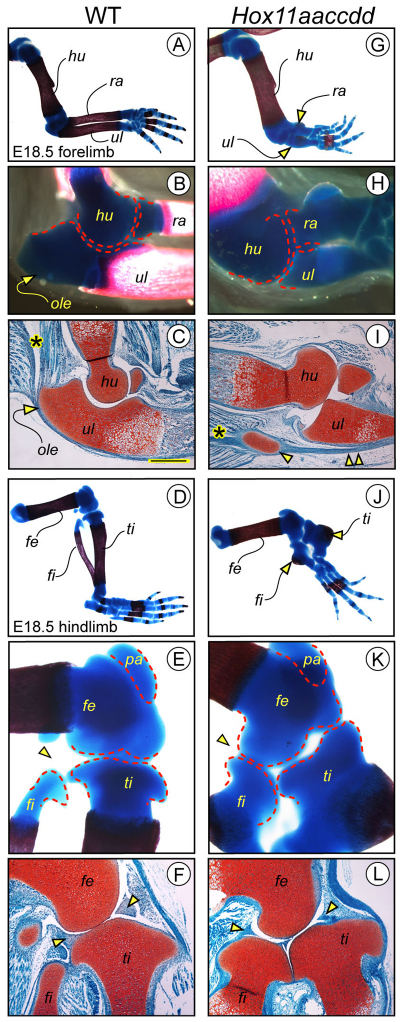
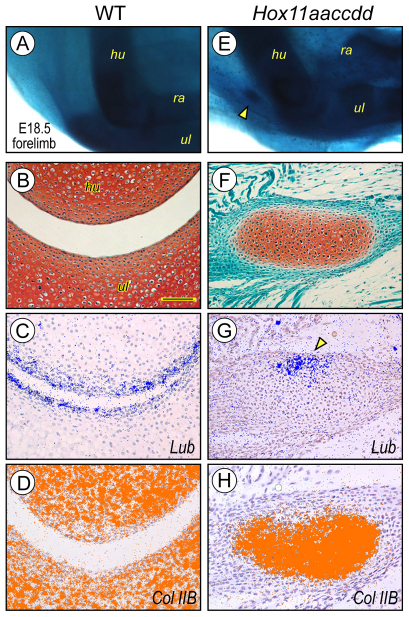
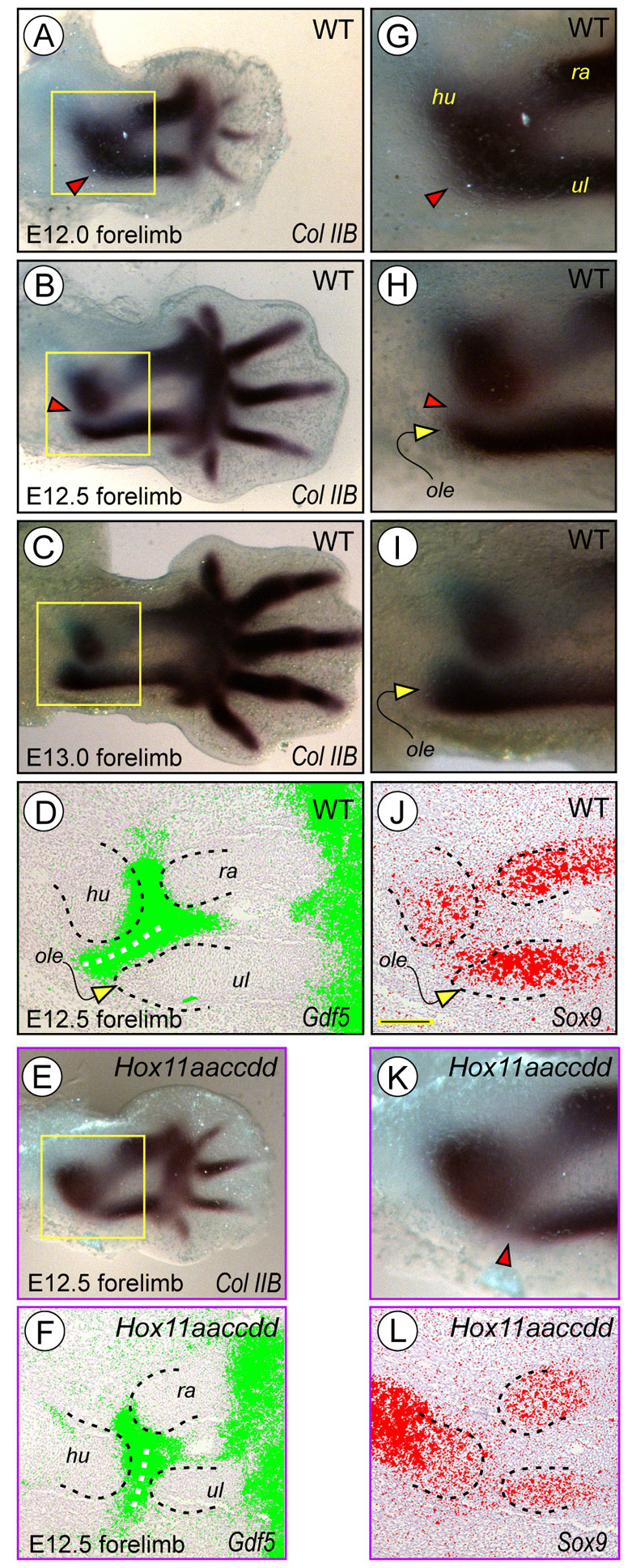
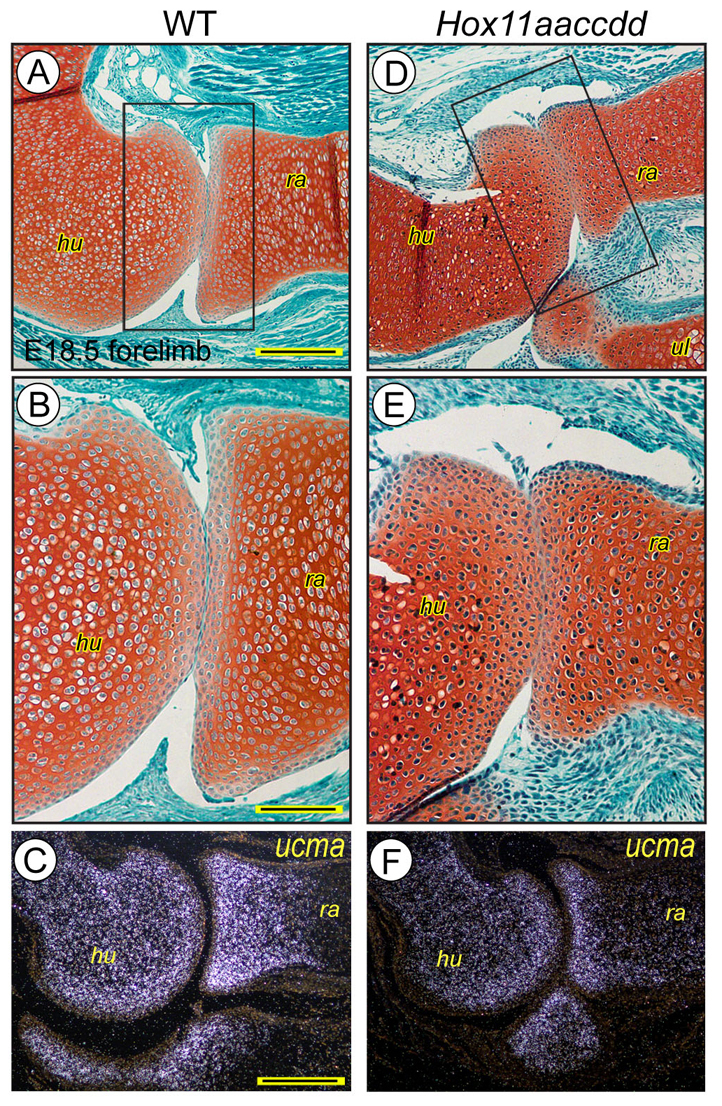
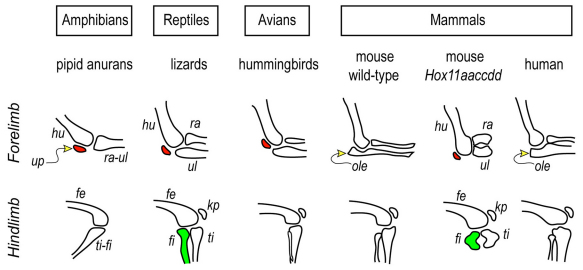
Hox11 genes are essential for zeugopod skeletal element development but their roles in synovial joint formation remain largely unknown. Here, we show that the elbow and knee joints of mouse embryos lacking all Hox11 paralogous genes are specifically remodeled and reorganized. The proximal ends of developing mutant ulna and radius elements became morphologically similar and formed an anatomically distinct elbow joint. The mutant ulna lacked the olecranon that normally attaches to the triceps brachii muscle tendon and connects the humerus to the ulna. In its place, an ulnar patella-like element developed that expressed lubricin on its ventral side facing the joint and was connected to the triceps muscle tendon. In mutant knees, both tibia and fibula fully articulated with an enlarged femoral epiphyseal end that accommodated both elements, and the neo-tripartite knee joint was enclosed in a single synovial cavity and displayed an additional anterior ligament. The mutant joints also exhibited a different organization of the superficial zone of articular cartilage that normally exerts an anti-friction function. In conclusion, Hox11 genes co-regulate and coordinate the development of zeugopod skeletal elements and adjacent elbow and knee joints, and dictate joint identity, morphogenesis and anatomical and functional organization. Notably, the ulnar patella and tripartite knee joints in the mouse mutants actually characterize several lower vertebrates, including certain reptiles and amphibians. The re-emergence of such anatomical structures suggests that their genetic blueprint is still present in the mouse genome but is normally modified to the needs of the mammalian joint-formation program by distinct Hox11 function.
Figures
References
-
- Archer C. W., Dowthwaite G. P., Francis-West P. (2003). Development of synovial joints. Birth Defects Research, Pt. C 69, 144-155 - PubMed
-
- Boulet A. M., Capecchi M. R. (2003). Multiple roles of Hoxa11 and Hoxd11 in the formation of the mammalian forelimb zeugopod. Development 131, 299-309 - PubMed
-
- Carroll S. B. (2008). Evo-Devo and an expanding evolutionary synthesis; a genetic theory of morphological evolution. Cell 134, 25-36 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
