

Renewed focus on the developing human neocortex
- PMID: 20979582
- PMCID: PMC2992407
- DOI: 10.1111/j.1469-7580.2010.01281.x
Renewed focus on the developing human neocortex
Abstract
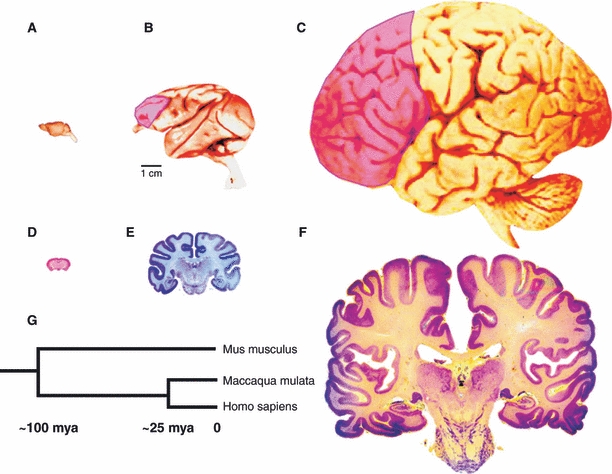
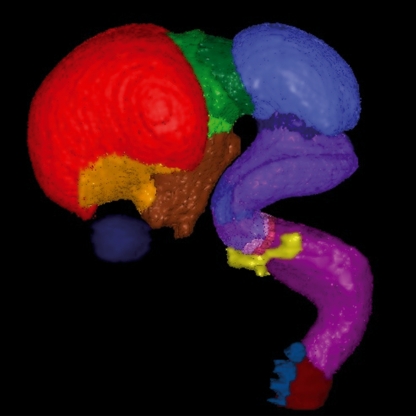
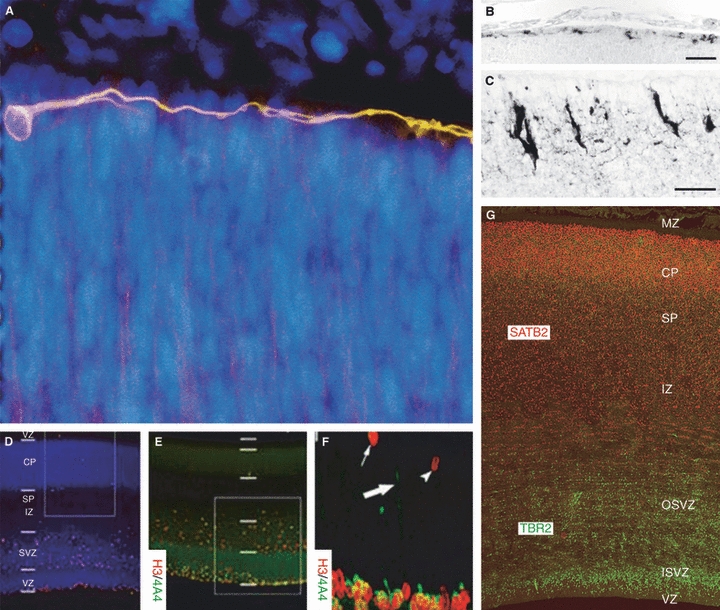
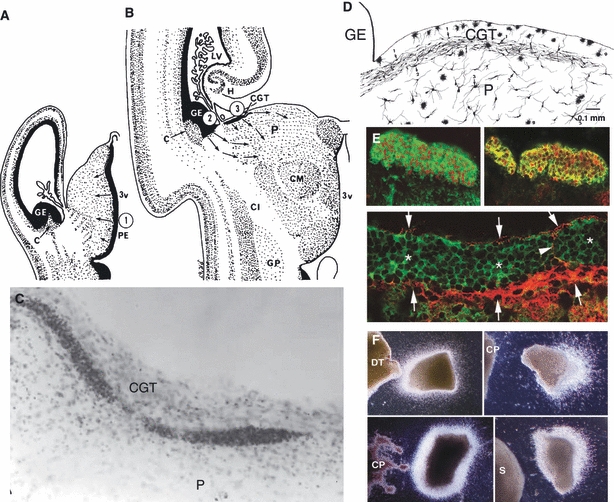
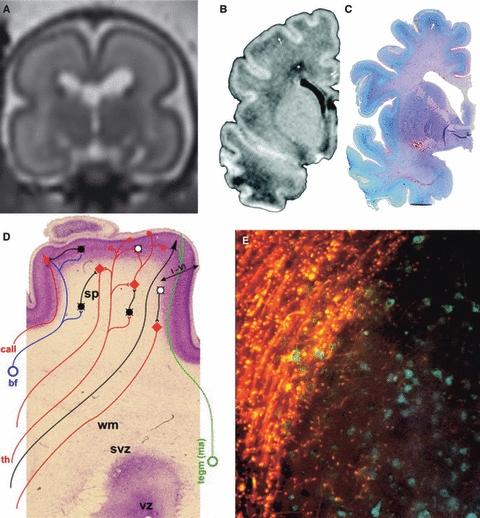
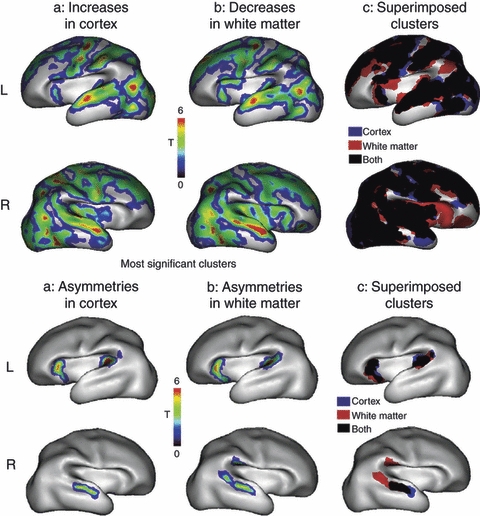
Many specifically human psychiatric and neurological conditions have developmental origins. Rodent models are extremely valuable for the investigation of brain development, but cannot provide insight into aspects that are specifically human. The human brain, and particularly the cerebral cortex, has some unique genetic, molecular, cellular and anatomical features, and these need to be further explored. Cortical expansion in human is not just quantitative; there are some novel types of neurons and cytoarchitectonic areas identified by their gene expression, connectivity and functions that do not exist in rodents. Recent research into human brain development has revealed more elaborated neurogenetic compartments, radial and tangential migration, transient cell layers in the subplate, and a greater diversity of early-generated neurons, including predecessor neurons. Recently there has been a renaissance of the study of human brain development because of these unique differences, made possible by the availability of new techniques. This review gives a flavour of the recent studies stemming from this renewed focus on the developing human brain.
© 2010 The Authors. Journal of Anatomy © 2010 Anatomical Society of Great Britain and Ireland.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
