

Decision threshold modulation in the human brain
- PMID: 20980586
- PMCID: PMC6634811
- DOI: 10.1523/JNEUROSCI.2371-10.2010
Decision threshold modulation in the human brain
Abstract
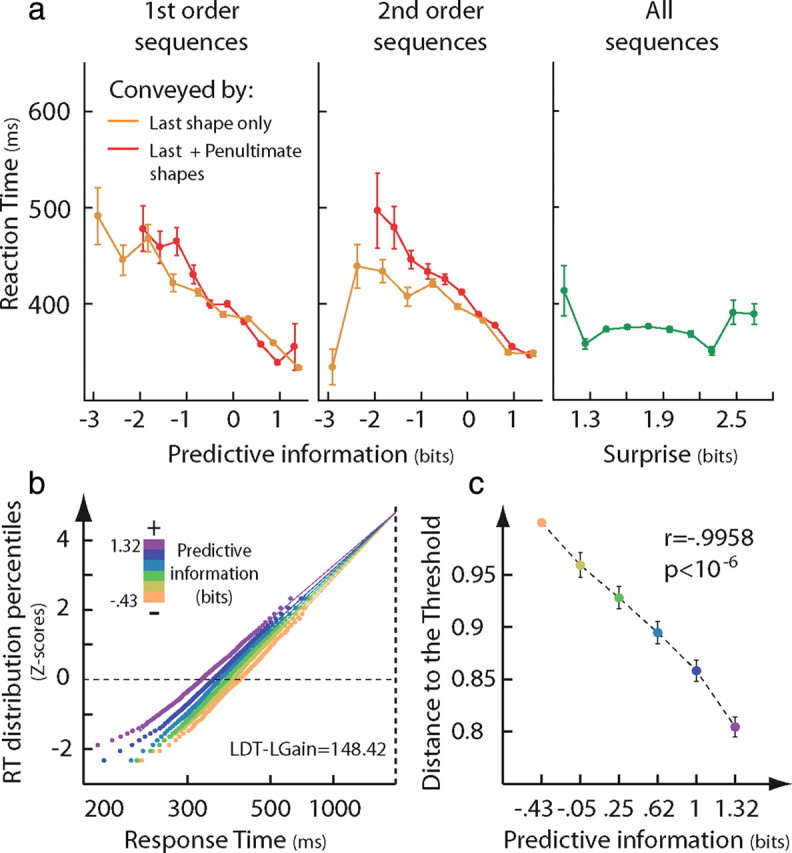
Perceptual decisions are made when sensory evidence accumulated over time reaches a decision threshold. Because decisions are also guided by prior information, one important factor that is likely to shape how a decision is adaptively tuned to its context is the predictability of forthcoming events. However, little is known about the mechanisms underlying this contextual regulation of the perceptual decision-making process. Mathematical models of decision making predict two possible mechanisms supporting this regulation: an adjustment of the distance to the decision threshold, which leads to a change in the amount of accumulated evidence required to make a decision, or a gain control of the sensory evidence, leading to a change in the slope of the sensory evidence accumulation. Here, we show that predictability of the forthcoming event reduces the distance to the threshold of the decision. Then, combining model-driven fMRI and the framework of information theory, we show that the anterior cingulate cortex (ACC) adjusts the distance to the decision threshold in proportion to the current amount of predictive information and that the dorsolateral cortex (DLPFC) codes the accumulation of sensory evidence. Moreover, the information flow from the ACC to the DLPFC region that accumulates sensory evidence increases when optimal adjustment of the distance to the threshold requires more complex computations, reflecting the increased weight of ACC's regulation signals in the decision process. Our results characterize the respective contributions of the ACC and the DLPFC to contextually optimized decision making.
Figures







References
-
- Behrens TE, Woolrich MW, Walton ME, Rushworth MF. Learning the value of information in an uncertain world. Nat Neurosci. 2007;10:1214–1221. - PubMed
-
- Berlyne DE. Uncertainty and conflict: a point of contact between information-theory and behavior-theory concepts. Psychol Rev. 1957;64:329–339. - PubMed
-
- Bogacz R. Optimal decision network with distributed representation. Neural Netw. 2007a;20:564–576. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources