

Molecular mechanisms of the whole DNA repair system: a comparison of bacterial and eukaryotic systems
- PMID: 20981145
- PMCID: PMC2957137
- DOI: 10.4061/2010/179594
Molecular mechanisms of the whole DNA repair system: a comparison of bacterial and eukaryotic systems
Abstract
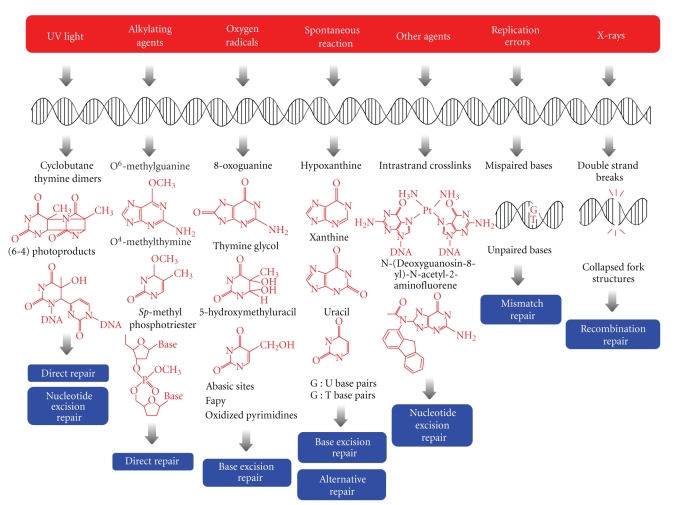
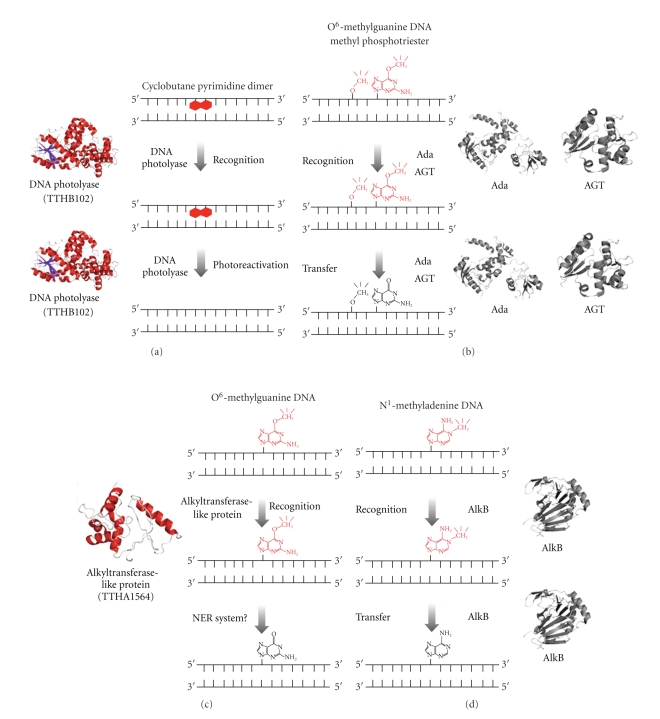
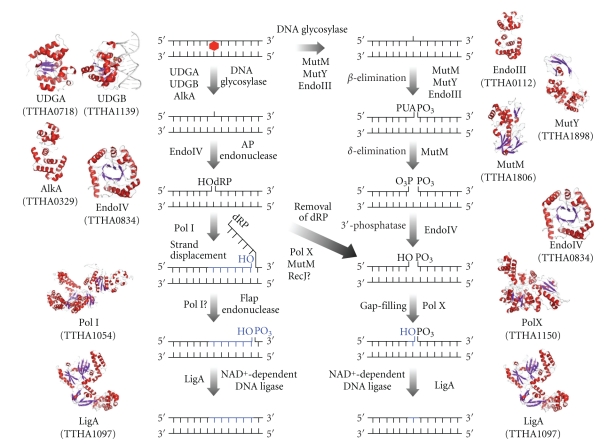
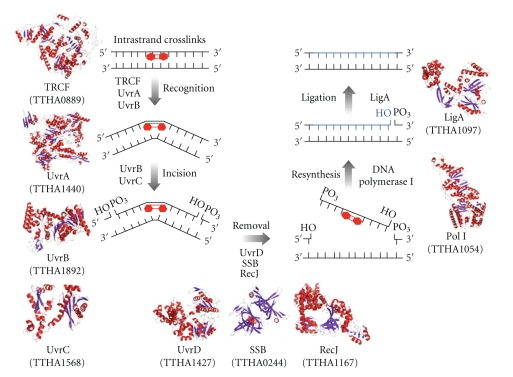
DNA is subjected to many endogenous and exogenous damages. All organisms have developed a complex network of DNA repair mechanisms. A variety of different DNA repair pathways have been reported: direct reversal, base excision repair, nucleotide excision repair, mismatch repair, and recombination repair pathways. Recent studies of the fundamental mechanisms for DNA repair processes have revealed a complexity beyond that initially expected, with inter- and intrapathway complementation as well as functional interactions between proteins involved in repair pathways. In this paper we give a broad overview of the whole DNA repair system and focus on the molecular basis of the repair machineries, particularly in Thermus thermophilus HB8.
Figures

References
-
- Schärer OD. Chemistry and biology of DNA repair. Angewandte Chemie. 2003;42(26):2946–2974. - PubMed
-
- Altieri F, Grillo C, Maceroni M, Chichiarelli S. DNA damage and repair: from molecular mechanisms to health implications. Antioxidants and Redox Signaling. 2008;10(5):891–937. - PubMed
-
- Friedberg EC, Walker GC, Siede W. DNA Repair and Mutagenesis. Washington, DC, USA: ASM Press; 2006.
-
- Oshima T, Imahori K. Description of Thermus thermophilus (Yoshida and Oshima) comb. nov., a nonsporulating thermophilic bacterium from a Japanese thermal spa. International Journal of Systematic Bacteriology. 1974;24(1):102–112.
LinkOut - more resources
Full Text Sources
Other Literature Sources
