

Controlling Ca2+-activated K+ channels with models of Ca2+ buffering in Purkinje cells
- PMID: 20981513
- PMCID: PMC3411306
- DOI: 10.1007/s12311-010-0224-3
Controlling Ca2+-activated K+ channels with models of Ca2+ buffering in Purkinje cells
Abstract
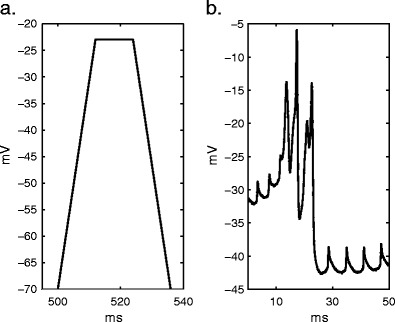
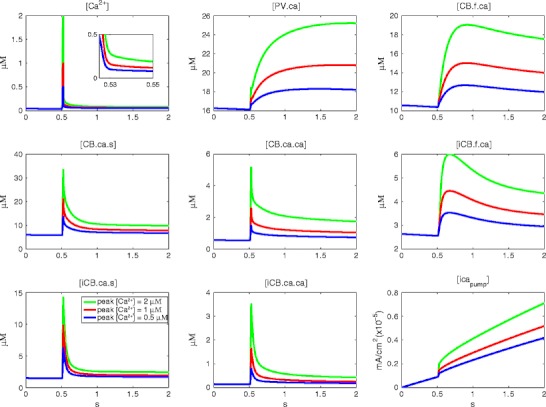
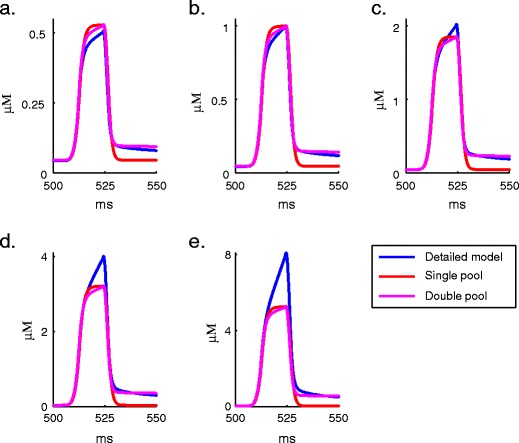
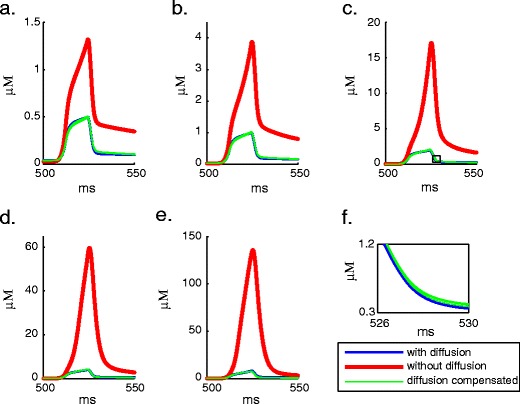
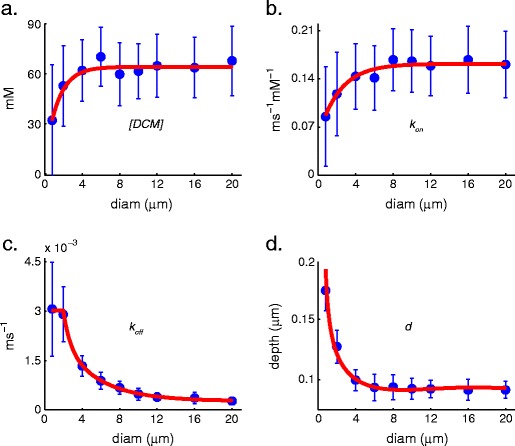
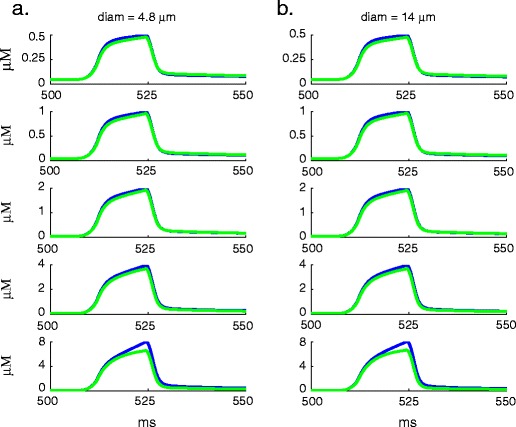
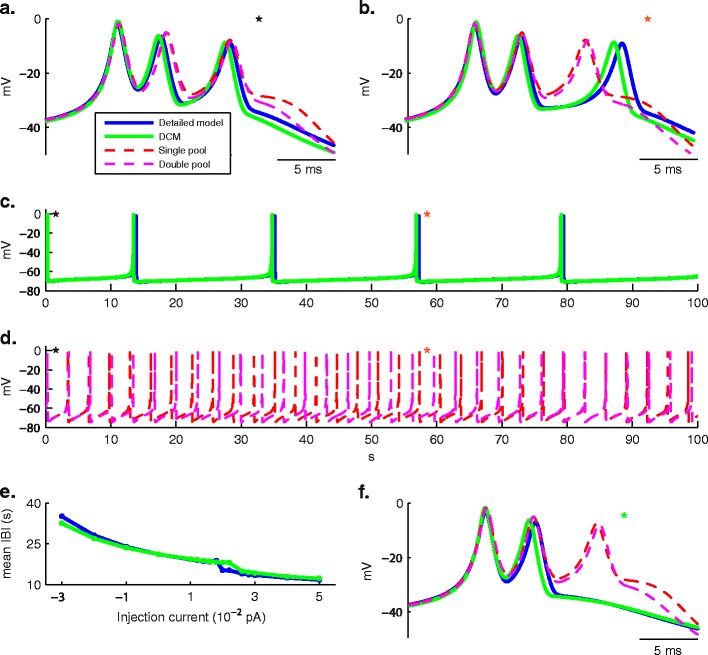
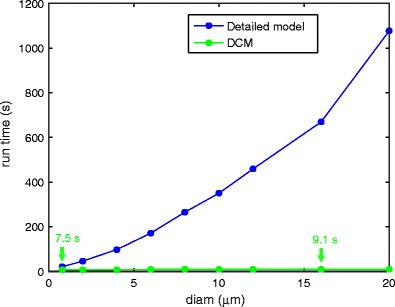
Intracellular Ca(2+) concentrations play a crucial role in the physiological interaction between Ca(2+) channels and Ca(2+)-activated K(+) channels. The commonly used model, a Ca(2+) pool with a short relaxation time, fails to simulate interactions occurring at multiple time scales. On the other hand, detailed computational models including various Ca(2+) buffers and pumps can result in large computational cost due to radial diffusion in large compartments, which may be undesirable when simulating morphologically detailed Purkinje cell models. We present a method using a compensating mechanism to replace radial diffusion and compared the dynamics of different Ca(2+) buffering models during generation of a dendritic Ca(2+) spike in a single compartment model of a PC dendritic segment with Ca(2+) channels of P- and T-type and Ca(2+)-activated K(+) channels of BK- and SK-type. The Ca(2+) dynamics models used are (1) a single Ca(2+) pool; (2) two Ca(2+) pools, respectively, for the fast and slow transients; (3) detailed Ca(2+) dynamics with buffers, pump, and diffusion; and (4) detailed Ca(2+) dynamics with buffers, pump, and diffusion compensation. Our results show that detailed Ca(2+) dynamics models have significantly better control over Ca(2+)-activated K(+) channels and lead to physiologically more realistic simulations of Ca(2+) spikes and bursting. Furthermore, the compensating mechanism largely eliminates the effect of removing diffusion from the model on Ca(2+) dynamics over multiple time scales.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
