

Transcription of the T4 late genes
- PMID: 21029432
- PMCID: PMC2988020
- DOI: 10.1186/1743-422X-7-288
Transcription of the T4 late genes
Abstract
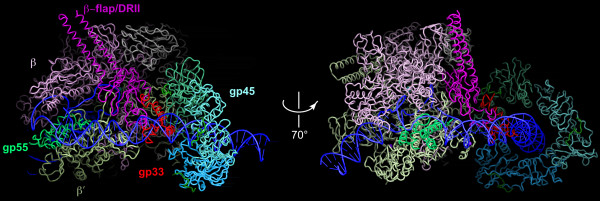
This article reviews the current state of understanding of the regulated transcription of the bacteriophage T4 late genes, with a focus on the underlying biochemical mechanisms, which turn out to be unique to the T4-related family of phages or significantly different from other bacterial systems. The activator of T4 late transcription is the gene 45 protein (gp45), the sliding clamp of the T4 replisome. Gp45 becomes topologically linked to DNA through the action of its clamp-loader, but it is not site-specifically DNA-bound, as other transcriptional activators are. Gp45 facilitates RNA polymerase recruitment to late promoters by interacting with two phage-encoded polymerase subunits: gp33, the co-activator of T4 late transcription; and gp55, the T4 late promoter recognition protein. The emphasis of this account is on the sites and mechanisms of actions of these three proteins, and on their roles in the formation of transcription-ready open T4 late promoter complexes.
Figures

Similar articles
-
Dissection of the bacteriophage T4 late promoter complex.J Mol Biol. 2008 Jun 6;379(3):402-13. doi: 10.1016/j.jmb.2008.03.071. Epub 2008 Apr 7. J Mol Biol. 2008. PMID: 18455735 Free PMC article.
-
Abortive initiation of transcription at a hybrid promoter. An analysis of the sliding clamp activator of bacteriophage T4 late transcription, and a comparison of the sigma70 and T4 gp55 promoter recognition proteins.J Biol Chem. 1998 Dec 18;273(51):34042-8. doi: 10.1074/jbc.273.51.34042. J Biol Chem. 1998. PMID: 9852060
-
Activator-sigma interaction: A hydrophobic segment mediates the interaction of a sigma family promoter recognition protein with a sliding clamp transcription activator.J Mol Biol. 1998 Nov 27;284(2):195-203. doi: 10.1006/jmbi.1998.2166. J Mol Biol. 1998. PMID: 9813112
-
Old phage, new insights: two recently recognized mechanisms of transcriptional regulation in bacteriophage T4 development.FEMS Microbiol Lett. 1995 Apr 15;128(1):1-8. doi: 10.1111/j.1574-6968.1995.tb07491.x. FEMS Microbiol Lett. 1995. PMID: 7744235 Review.
-
Transcriptional control in the prereplicative phase of T4 development.Virol J. 2010 Oct 28;7:289. doi: 10.1186/1743-422X-7-289. Virol J. 2010. PMID: 21029433 Free PMC article. Review.
Cited by
-
A Shigella boydii bacteriophage which resembles Salmonella phage ViI.Virol J. 2011 May 19;8:242. doi: 10.1186/1743-422X-8-242. Virol J. 2011. PMID: 21595934 Free PMC article.
-
Sustained in situ protein production and release in the mammalian gut by an engineered bacteriophage.Nat Biotechnol. 2025 Feb 18:10.1038/s41587-025-02570-7. doi: 10.1038/s41587-025-02570-7. Online ahead of print. Nat Biotechnol. 2025. PMID: 39966654 Free PMC article.
-
From Processivity to Genome Maintenance: The Many Roles of Sliding Clamps.Genes (Basel). 2022 Nov 7;13(11):2058. doi: 10.3390/genes13112058. Genes (Basel). 2022. PMID: 36360296 Free PMC article. Review.
-
The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly.Viruses. 2022 Mar 28;14(4):700. doi: 10.3390/v14040700. Viruses. 2022. PMID: 35458430 Free PMC article. Review.
-
An introduction to transcription and gene regulation.J Biol Chem. 2010 Aug 20;285(34):25885-92. doi: 10.1074/jbc.X110.143867. Epub 2010 May 28. J Biol Chem. 2010. PMID: 20511220 Free PMC article. No abstract available.
References
-
- Karam JD, Editor-in-Chief. Molecular biology of bacteriophage T4. Washington, DC: American Society for Microbiology; 1994.
-
- Kassavetis GA, Zentner PG, Geiduschek EP. Transcription at bacteriophage T4 variant late promoters. An application of a newly devised promoter-mapping method involving RNA chain retraction. J Biol Chem. 1986;261:14256–14265. - PubMed
-
- Williams KP, Kassavetis GA, Herendeen DR, Geiduschek EP. In: Molecular Biology of Bacteriophage T4. Karam JD, editor. Washington, D.C.: American Society for Microbiology; 1994. Regulation of late-gene expression; pp. 161–175.
