

Iron loading-induced aggregation and reduction of iron incorporation in heteropolymeric ferritin containing a mutant light chain that causes neurodegeneration
- PMID: 21029774
- PMCID: PMC3040265
- DOI: 10.1016/j.bbadis.2010.10.010
Iron loading-induced aggregation and reduction of iron incorporation in heteropolymeric ferritin containing a mutant light chain that causes neurodegeneration
Abstract
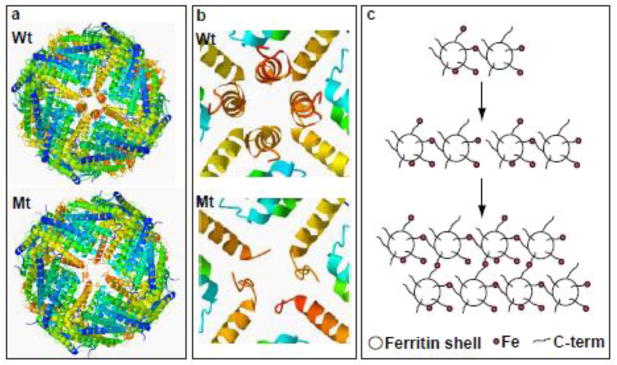
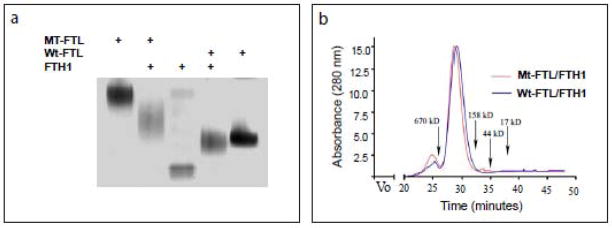
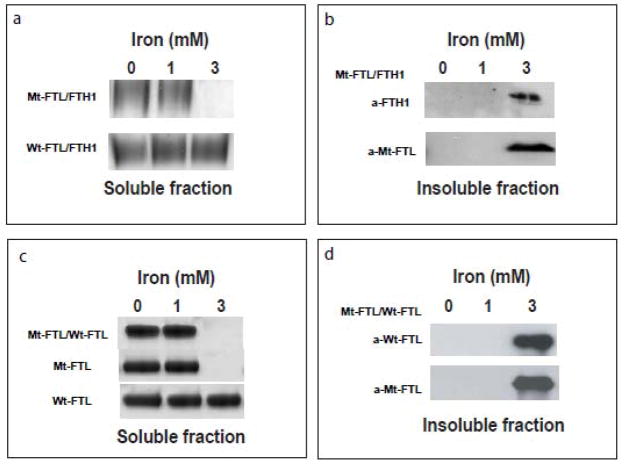
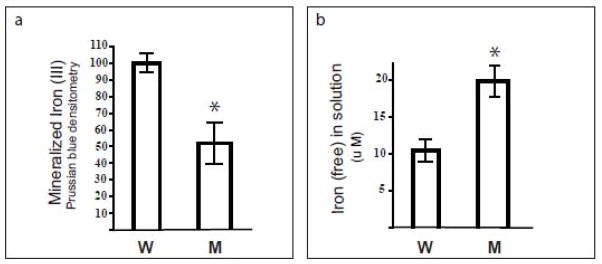
Hereditary ferritinopathy (HF) is a neurodegenerative disease characterized by intracellular ferritin inclusion bodies (IBs) and iron accumulation throughout the central nervous system. Ferritin IBs are composed of mutant ferritin light chain as well as wild-type light (Wt-FTL) and heavy chain (FTH1) polypeptides. In vitro studies have shown that the mutant light chain polypeptide p.Phe167SerfsX26 (Mt-FTL) forms soluble ferritin 24-mer homopolymers having a specific structural disruption that explains its functional problems of reduced ability to incorporate iron and aggregation during iron loading. However, because ferritins are usually 24-mer heteropolymers and all three polypeptides are found in IBs, we investigated the properties of Mt-FTL/FTH1 and Mt-FTL/Wt-FTL heteropolymeric ferritins. We show here the facile assembly of Mt-FTL and FTH1 subunits into soluble ferritin heteropolymers, but their ability to incorporate iron was significantly reduced relative to Wt-FTL/FTH1 heteropolymers. In addition, Mt-FTL/FTH1 heteropolymers formed aggregates during iron loading, contrasting Wt-FTL/FTH1 heteropolymers and similar to what was seen for Mt-FTL homopolymers. The resulting precipitate contained both Mt-FTL and FTH1 polypeptides as do ferritin IBs in patients with HF. The presence of Mt-FTL subunits in Mt-FTL/Wt-FTL heteropolymers also caused iron loading-induced aggregation relative to Wt-FTL homopolymers, with the precipitate containing Mt- and Wt-FTL polypeptides again paralleling HF. Our data demonstrate that co-assembly with wild-type subunits does not circumvent the functional problems caused by mutant subunits. Furthermore, the functional problems characterized here in heteropolymers that contain mutant subunits parallel those problems previously reported in homopolymers composed exclusively of mutant subunits, which strongly suggests that the structural disruption characterized previously in Mt-FTL homopolymers occurs in a similar manner and to a significant extent in both Mt-FTL/FTH1 and Mt-FTL/Wt-FTL heteropolymers.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
References
-
- Zecca L, Youdim MB, Riederer P, Connor JR, Crichton RR. Iron, brain ageing and neurodegenerative disorders. Nat Rev Neurosci. 2004;5:863–873. - PubMed
-
- Crichton R. Iron metabolism from molecular mechanisms to clinical consequences. 3. Wiley; West Sussex, U.K: 2009.
-
- Stefanini S, Cavallo S, Wang CQ, Tataseo P, Vecchini P, Giartosio A, Chiancone E. Thermal stability of horse spleen apoferritin and human recombinant H apoferritin. Arch Biochem Biophys. 1996;325:58–4. - PubMed
-
- Santambrogio P, Levi S, Arosio P, Palagi L, Vecchio G, Lawson DM, Yewdall SJ, Artymiuk PJ, Harrison PM, Jappelli R, Cesareni G. Evidence that a salt bridge in the light chain contributes to the physical stability difference between heavy and light human ferritins. J Biol Chem. 1992;267:14077–14083. - PubMed
-
- Levi S, Santambrogio P, Cozzi A, Rovida E, Corsi B, Tamborini E, Spada S, Albertini A, Arosio P. The role of the L-chain in ferritin iron incorporation. Studies of homo and heteropolymers. J Mol Biol. 1994;238:649–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
